Страница не найдена
Золотой век Голливуда раскрыл целую плеяду талантливых актрис, вошедших в историю кинематографа. Но лишь единицы известны всему миру, одна из таких ярчайших звезд — Одри Хепберн. Ее называют иконой стиля, лаконичная элегантность была основой гардероба. Не меньшего восхищения заслуживает и внешность актрисы. Яркие, знойные красотки меркнут в сравнении с утонченной аристократкой Хепберн. Особый магнетизм заставлял забывать о неидеальности черт лица, актриса создала новые стандарты красоты. Сумела найти свой индивидуальный стиль, который сегодня наследуют миллионы.
Прическа Одри Хепберн всегда отличалась аккуратной формой, безупречные линии без намека на небрежность, легкомысленность. Сдержанность, скрупулезность к деталям, строгое воспитание не позволяли появиться на людях без идеальной укладки и безупречного макияжа. Актриса прекрасно знала, как влияет на окружающих, интуитивно выбирала лучшие фильмы, роли, платья, стрижки. Она стала законодательницей современной моды, подарив миру маленькое черное платье, фактурные очки «кошачий глаз», мейкап с выразительными бровями. Стройная, утонченная звезда создавала собственные тенденции, подчеркивающие ее индивидуальность. Предпочитала высокие прически с собранными локонами, открывающие грациозный силуэт, лебединую шею. Великолепно выглядела в одежде любого стиля, но ее стрижки и укладки относились к классическим моделям.
Природные внешние данные
Секрет успеха Одри Хепберн заключается в ее органичности, никогда не шла на поводу у капризной моды. Легкая, грациозная, с огромными томными глазами, она завораживала с первого взгляда. В этот период Голливудом правят легкомысленные блондинки, жизнерадостные, раскрепощенные, невероятно привлекательные.
Актриса была полной противоположностью, утонченная, сдержанная, не привыкшая проявлять бурные эмоции, отличалась и внешними данными. Миндалевидные глаза, окутанные грустью, пышные ресницы, густые, выразительные брови, аккуратная стрижка средней длины. Все очень просто, но невероятно гармонично, на протяжении всей карьеры сохраняла природную красоту.
Все очень просто, но невероятно гармонично, на протяжении всей карьеры сохраняла природную красоту.
Темно-каштановые волосы укладывала в собранные высокие прически. Но и варианты с распущенными локонами не выглядели легкомысленно, вызывающе. Каждой, пряди, каждому завитку уделяется внимание, симметрия линий подчеркивает аристократическое происхождение, безупречный вкус, тонкое ощущение моды.
Основные принципы выбора прически:
- обладает густыми локонами прямой структуры темно-каштанового цвета;
- редко прибегала к окрашиваниям, поэтому запомнилась зрителям исключительно прическами с темными волосами;
- укладки максимально открывают лицо, периодически создавала короткие челки, не достигающих линии бровей;
- минимальный объем, используемых укладочных средств, позволял создать аккуратную форму, при этом сохранялась естественность линий, не было намека на искусственность;
- шпильки, невидимки помогали создавать классические прически;
- предпочитала золотую середину при выборе фактуры волос, не использовала футуристические образы с прямыми прядями и мелкие игривые кудряшки, легкая волна с плавными изгибами подчеркивала утонченность, женственность актрисы.
Какая прическа была в детстве, юности
Будущая звезда родилась в Брюсселе, по линии матери род состоял из аристократов, военных высших чинов, придворных, государственных служащих. Ее бабушка была баронесса, дедушка — высший чиновник в министерстве юстиции, отец работал почетным консулом. Девочка росла в роскоши, богатстве, с детства прививались правила хорошего тона. Но все изменила Вторая мировая война, голод привел к развитию анемии и проблемам со здоровьем.
Несмотря на все тяготы и лишения, по воспоминаниям самой актрисы, детство у нее было счастливое, светлые периоды затмевали страх, постоянную тревогу. Ее стрижка соответствовала военному периоду, короткое каре с челкой позволяло минимизировать уход за детскими волосами. Темные локоны всегда отличались густотой, даже простая стрижка подчеркивала миловидность малышки.
Ее стрижка соответствовала военному периоду, короткое каре с челкой позволяло минимизировать уход за детскими волосами. Темные локоны всегда отличались густотой, даже простая стрижка подчеркивала миловидность малышки.
Юная Одри работала медсестрой и параллельно брала уроки балета у прославленных танцоров. Ей пророчили блестящее будущее, но голод, перенесенный в военные годы, отразился на здоровье девушки. Недолго думая, решает попробовать себя в актерстве. Яркие внешние данные, утонченные манеры, грациозность балерины позволили быстро завоевывать расположение не только режиссеров, но и всей съемочной группы.
Ее прически отличаются легкостью, присущей юности. Средняя длина, завитые локоны, короткая челка — основные элементы, присутствующие и после присвоения статуса иконы стиля. Пройдет несколько лет и укладки будут идеальны, продуманы до мелочей. А пока Одри может быть юной, беспечной, скромной, романтичной, прекрасной, не догадываясь, какие подарки судьба ей приготовила.
Прически звезды
Культовые прически Одри Хепберн не теряют актуальности сегодня. Короткую стрижку «Пикси» выбирают барышни подобного типажа внешности — хрупкие, утонченные. Высокая укладка с диадемой стала любимой моделью звезд для выхода на красную дорожку, создают ее и для особых торжеств, используя для образа невесты, выпускницы. Одри Хепберн стала примером безупречного вкуса, строгой элегантности.
50-е
Актриса начинает сниматься в эпизодах, юной красотке с внешностью королевской особы не долго пришлось быть на втором плане, режиссеры утверждают на главные роли. К картине «Римские каникулы» длинные локоны состригает по сюжету фильма, так появляется знаменитая «Пикси». Ее успех обусловлен легкостью, динамичностью формы и простотой в уходе. Стрижка, созданная для актрисы, отличается от современных андрогенных вариантов.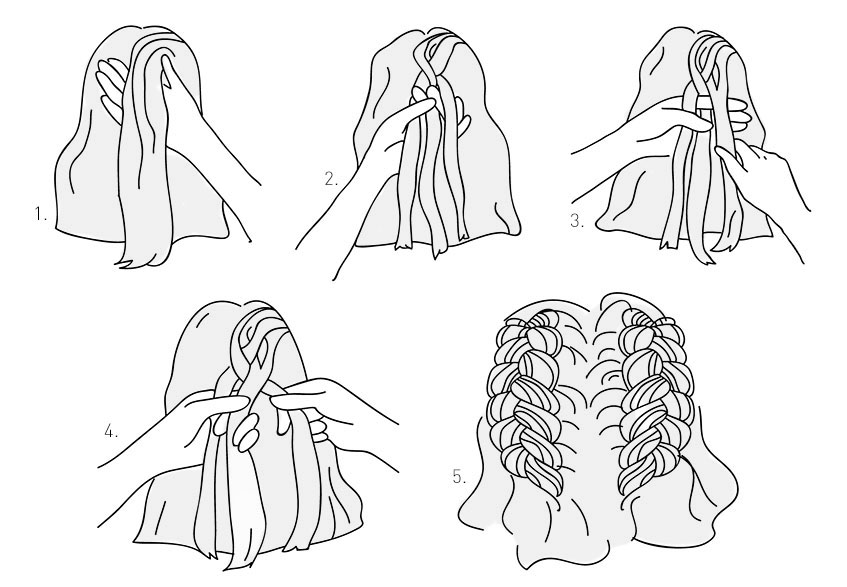 Пряди укладывает вверх или назад, отсутствуют резкие линии, подчеркивает романтичность натуры.
Пряди укладывает вверх или назад, отсутствуют резкие линии, подчеркивает романтичность натуры.
Дополнен образ густой челкой до середины лба, идеальный вариант, чтобы подчеркнуть выразительные черты лица и белоснежную кожу. Прическа выгодно подчеркивает внешность актрисы, ее точеные скулы, правильные черты. Универсальная стрижка позволяет быть легкой и беззаботной, а через мгновение восхищать утонченной элегантностью.
Не менее яркой является прическа на среднюю длину. Густые, темно-каштановые локоны завивает крупной волной, плавные изгибы подчеркивают огромные глаза. Индивидуальность образа позволило создать свой собственный стиль, который заметно отличался от послевоенных модных веяний. Сдержанная скромная укладка, для создания которой требовалось всего несколько невидимок, произвела настоящий фурор.
Обратите внимание! Актриса выбирала прически, в которых чувствовала себя органично, не следовала модным веяниям. Прямой пробор, густая челка и средняя длина темных локонов позволили создать незабываемый образ, успешно используемый и сегодня.
60-е
В начале 60-х выходит культовый фильм «Завтрак у Тиффани», кинолента стала знаковой в карьере актрисы. Роль легкомысленной охотницы за богатым мужем далась не просто сдержанной, утонченной Одри. Но именно благодаря этой героини было создано легендарное маленькое черное платье в соавторстве с Юбером Живанши. Также классикой парикмахерского искусства стала укладка «Ракушка». Волосы зачесываются назад и закручиваются по спирали, роскошная вечерняя прическа дополнена легкой короткой челкой.
Также для участия в съемках пришлось высветлить отдельные пряди, мелирование подчеркнуло плавные завитки праздничной укладки. Образ стал эталоном женственности и элегантности, в современном мире не теряет актуальности.
Великолепно выглядит со стрижкой каре, в отличие от детской прически, предпочитает удлиненный вариант. Не использует челку, зачесывает локоны назад, фиксацией служат ленты и широкие ободки. Срезы прядей укладывает ровно или кончиками наружу. Прическа подчеркивает романтичность, легкость актрисы, позволяя выглядеть сногсшибательно не только на красной дорожке, но и в повседневной жизни.
В середине 60–х появляется с новой стрижкой, боб с высоким начесом позволяет выглядеть по-другому, при этом сохраняются узнаваемый стиль Одри. Популярные модели отличались объемной макушкой, напоминающей форму шара. Достигался подобный эффект за счет начеса и применения стайлинга. Новым в образе актрисы стала густая челка на одну сторону, дополненная фактурными прядями у висков.
Стильная стрижка в сочетании с модными стрелками стали основой модных тенденций 60-х. Сдержанная сексуальность Хепберн быстро сделала ее кумиром. Укладки и наряды копировали девушки по всему миру, не понимая, что Одри выбирает лишь те варианты, которые подчеркивать ее индивидуальность.
70-е
Актриса не перестает экспериментировать, ее новая стрижка — «Аврора» на короткие волосы. Многоуровневая модель создает эффект объемной копны, особенно если пряди уложены завитками. На макушке и темени локоны стригутся коротко, на затылке сохраняется удлинение. К привычной сдержанной элегантности добавляет нотки креативности, ведь стрижка была популярна среди неформальных движений.
Модель обладает корректирующими свойствами, делает моложе, позволяет выделить скульптурные скулы. Благодаря завиткам создается женственный, романтичный образ, универсальная модель отлично сочетается с любимыми классическими нарядами актриса. После фильма «Завтрак у Тиффани» актриса не меняла цвет волос, отдавая предпочтение натуральному оттенку.
80-е
Знаменитость вновь создает новую прическу, на этот раз отдает предпочтение лаконичному пучку, уложенному на затылке. Подобный вариант могут позволить себе лишь стройные, изящные женщины с идеальной формой лица, как у Одри Хепберн. Актриса является примером как можно стареть красиво. Сохраняет женственность и грациозность, безупречный вкус и внимание к деталям позволяет оставаться кумиром миллионов.
Прическа полностью открывает лицо, небольшой объем области темени подчеркивает четкие линии овала. Тогда как дамы бальзаковского возраста выбирают многослойные стрижки с челкой в надежде выглядеть моложе, Одри Хепберн, наоборот, достойно принимает неизбежность увядания.
90-е
В последние годы своей жизни актриса не снимается в кино, но ведет активную деятельность в ЮНИСЕФ. Знаменитость остается верной скромной укладке на среднюю длину волос. Универсальный образ позволяет выглядеть элегантно как в повседневности, также подходит для особых случаев. Высокая укладка подчеркивает изящность, которую актриса сохранила до конца своих дней.
Самая известная прическа звезды
Актриса создала немало ярких образов на экране, но наиболее запоминающейся стала роль в ленте «Завтрак у Тиффани». Сама звезда тщательно прорабатывала каждую деталь прически, макияжа, наряда. Неудивительно, что и в повседневности Одри Хепберн выглядела также сногсшибательно, как и ее героиня Холли Голайтли. В ее кинокарьере преобладали романтические комедии, актриса и в жизни отличалась жизнерадостностью, но ее сложно назвать легкомысленной особой. Стильная, сдержанная, элегантная прическа «Ракушка» стала знаковой в образах звезды.
Собранные локоны создают гармоничную форму, включающую женственность и простоту. Укладка в виде завитка подчеркивает хрупкий силуэт и кукольное лицо с огромными глубокими глазами. Высокая прическа делает выше, акцентирует внимание на тонкой шее, линии плеч. Стала бестселлером среди вечерних причесок, кажется, образ Одри Хепберн примерили абсолютно все звезды кино и шоу-бизнеса.
Высокая прическа делает выше, акцентирует внимание на тонкой шее, линии плеч. Стала бестселлером среди вечерних причесок, кажется, образ Одри Хепберн примерили абсолютно все звезды кино и шоу-бизнеса.
Стильный вариант подходит не только для особых случаев, уместен и для повседневности. Густая челка средней длины придает легкость, свободу, но в то же время отсутствует небрежность, локоны сохраняют аккуратную форму. Подобные укладки актриса сочетала с мягкими трикотажными свитерами, брючными комплектами, идеально подходили для маленького черного платья.
Образ принцессы сегодня часто используют невесты, выпускницы. Актриса уложила пряди в высокую ракушку, а свободные кончики завила легкой волной. Украшением служат диадемы, моментально превращающие простую прическу, в укладку достойную самой королевы. Короткая густая челка а-ля Наташа Ростова подчеркивает выразительные черты, безупречный овал.
Создать прическу как у звезды можно самостоятельно в домашних условиях. Выполняется на среднюю и длину ниже плеч, для придания густоты можно накрутить пряди. Чтобы прическа держала форму, нужно обработать волосы пенкой или муссом. Не стоит создавать тугой завиток, это лишит прическу естественного объема. На затылке фиксируется невидимками и шпильками.
Кому подходит:
- прическу рекомендуется выбирать барышням с треугольным или правильным овалом, максимально открывает лицо, сложно скорректировать недостатки;
- можно сочетать с челкой или спрятать ее, зачесав с основной копной назад;
- лучше смотрится с монохромным окрашиванием, но как показал пример актрисы, мелирование придает индивидуальность прическе;
- сегодня используется при создании образа невесты, выпускницы или для вечернего торжества, но при отсутствии украшений подходит и для делового, повседневного стиля.


]]>
Два колоска по бокам наизнанку. Инструкция плетения колоска самой себе
Косичка «Колосок» является одной из самых популярных независимо от возраста. Эта прическа является актуальной как для повседневности, так и для различных торжественных мероприятий. Колосок подойдет для волос различной длинны, а плести такую косу совсем не сложно. Из нашей статьи вы узнаете о различных способах плетения колоска.
Плюсы и минусы прически
Ее достоинствами являются:
- комфортность – волосы не распадаются и не лезут в глаза;
- универсальность – эта прическа уместна будет как в офисе, повседневной жизни, так и в торжественных случаях;
- простота исполнения – такую косу можно заплести себе самостоятельно.
Данная прическа имеет и недостатки:
- обладательницам вьющихся локонов необходимо будет их выпрямить, для того чтобы заплести;
- также придется увлажнять и гладкие рассыпающиеся волосы, так как коса будет распадаться;
- трудно удерживать основную косу и одновременно подхватывать боковые пряди.
Но чтобы не возникало трудностей в плетении, необходимо немного потренироваться.
Пошаговая инструкция плетения себе
Любую прическу сделать самой себе проблематично. Но можно упростить процесс, сев между двух зеркал так, чтобы видно было заднюю часть головы и руки.
Теперь следует начинать процесс плетения:
- Отделить часть волос на макушке.
- Разделить ее на три пряди.
- Первую прядку, крайнюю с левой стороны, пропустить между двумя остальными.
- Затем проделать то же с правой стороны.
- Теперь незадействованную прядь располагаем между остальными. При этом, прихватывая с левой стороны небольшую часть свободных волос и добавляя в плетение.
- То же проделывается и с правой стороны.
- Таким же образом колосок плетется до шеи или в зависимости от длины шевелюры.
- Дальше заплетается обычная коса, а кончики собираются резинкой.
Чтобы прическа выглядела опрятно и красиво, необходимо захватывать одинаковые по толщине пряди с двух сторон.
На бок
Интересен и способ плетения колоска на бок, а не посередине. Для этого необходимо:
- Сделать косой пробор.
- От большей части шевелюры с правой стороны отделить небольшую прядь шириной приблизительно 5 см и разделить ее на три секции.
- Сначала плетется обычная коса.
- На третьем шаге вплетаются боковые пряди, которые захватываются поочередно, то справа, то слева.
- Дойдя до уровня шеи повернуть плетение влево.
- Доплести до левого уха, собрать все волосы и продолжать плести обычную косу в зависимости от их длины.
- Свободные кончики собрать резинкой.
Так как этот вид колоска больше подвержен распадению, то перед началом плетения можно на волосы нанести немного фиксирующего средства.
Вокруг головы
Такой вид прически собирает полностью всю волосяную массу и плетется следующим образом:
- Делается круговой пробор от средины лба и до затылочной части.
- Определяется центральная точка прически, для этого отмеряются равные промежутки от лобной и шейной части головы до макушки.
- От центральной точки лба начинается плетение колоска, но прядки подхватываются только с наружной стороны спирали.
- В конце последнего ряда, дойдя до необходимого места, плести обычную косичку.
- Собрать концы резиночкой и спрятать под нижним кругом косы, провести под ней насколько хватит длины, а потом спрятать.
Расплетая такую косу, получаются красивые и стойкие локоны, практически новая готовая прическа.
Вывернутый колосок
Способ плетения такого вида прически имеет следующую последовательность:
- Выбрать прядь вверху головы и разделить на три части.
- Захватить первый с левой стороны локон и завести его вниз под две остальные.
- Третью прядку подвести под первую и вторую, получается плетение внутрь.
- Опять проделываются эти шаги, но уже справа и слева подбираются свободные пряди.
- Дальше плетется по этой же схеме до конца волос, концы фиксируются.

Чтобы коса получилась объемной, необходимо немножко вытянуть пряди.
Прическа 2 колоска
Это более усложненный вариант, чем плетение одной косы, так как плетется в два раза дольше. Но чтобы быть красивой, надо потрудиться и сделать следующие шаги:
- Разделить волосы ровным пробором на две одинаковые части.
- С одной стороны волосяную массу собрать в хвост и убрать.
- С другой стороны разделить прядь мысленно на 3 секции.
- Первую, с левой стороны прядку, заложить в середину между остальными двумя.
- Третью – переложить между второй и первой.
- Далее повторять шаги, но уже добавляя волосы с боков, формируя колосок.
- Доплести косичку до конца и стянуть резинкой.
- Проделать такие же действия с другой частью шевелюры.
Можно собрать косы в пучок, и получится элегантная прическа.
Особенности плетения колоска ребенку
Заплетая косы девочке, нужно учитывать некоторые нюансы, а именно:
- дети непоседливы, поэтому высидеть могут минут 15 и за это время необходимо успеть сделать прическу;
- нельзя туго заплетать волосы, потому что у ребенка может разболеться голова;
- если локоны непослушные, их можно смочить только водой – никаких гелей, лаков и муссов.
Поэтому необходимо выбирать простые виды плетения, которые быстро плетутся.
Пошаговая инструкция того, как за несколько минут заплести колосок ребенку:
- Сначала необходимо смочить волосы тепленькой водой и расчесать мягкой расческой.
- Выделить прядь, начиная ото лба и поделить ее на три секции.
- Сначала заплетается обычная коса, потом перекрещивается левая прядка с центральной.
- Дальше вплетаются пряди с правой, затем с левой стороны.
- Закончить плести можно в районе шеи и собрать оставшиеся волосы резинкой.

Если удалось сделать эту прическу, то можно переходить к ее усложненному виду. Например, заплести два колоска. Для этого необходимо:
- Поделить волосы на две пряди ровным пробором.
- Одну сторону не трогать, а с другой сплести колосок, как описано выше.
- Затем проделать то же с другой стороны.
Научившись плести эту прическу, следует попробовать более сложный вариант – заплести колосок вокруг головы:
- Отделить прядь возле уха.
- Заплетать косу по линии лба до следующего уха, приплетая свободные пряди.
- Плести вокруг голову.
- Оставшуюся часть волос заплести в обычную косичку и спрятать внутри колоска.
- Зафиксировать невидимками.
Используя одну и ту же технику плетения, но заплетая косу туго, захватывая тонкие прядки либо слабо, вплетая локоны крупнее, прическа будет выглядеть по-разному. Первый вариант подходит молодым леди, а второй небрежный – лучше будет смотреться на взрослых дамах.
Прическа колосок – нежная и элегантная прическа, плести которую должна уметь каждая современная девушка. Сегодня колосок используют как красивое дополнение торжественного, делового и повседневного образов. Для создания элегантной прически можно бесконечно экспериментировать с ее формой и плетением.
Сегодня мы поговорим о том, как самостоятельно сделать красивую французскую косу.
БЫСТРАЯ НАВИГАЦИЯ ПО СТАТЬЕ
Классический вариант
Классический вариант подходит и для тонких и для густых волос
. «Французская коса» станет прекрасным дополнением праздничного наряда. Рассмотрим технологию ее создания подробнее.
- Тщательно расчешите шевелюру. Разделите ее горизонтальным пробором на две части, нижнюю зафиксируйте заколкой или резинкой.
- Возьмите верхние волосы и разделите их на три одинаковые пряди.
- Крайние пряди возьмите в руки. Средняя при этом должна оставаться свободной.
- Правую часть положите на центральную, затем на новую прядь по центру положите левую. Начало колоска плетется аналогично обычной косе.
- Затем придерживая левой рукой левую и центральную пряди, правой рукой захватите небольшую часть волос из нижней зоны и соедините ее с основной правой прядью. Подробную фото- и видео-инструкцию плетения вы можете увидеть ниже.
- Затем правую прядь переплетите со средней.
- Повторите шаг №5 с левой стороны.
- Повторяйте шаг №5 и шаг №7 до тех пор, пока плетение не дойдет до конца зоны роста волос.
- Зафиксируйте результат резинкой. Французская коса готова! Подробная технология ее создания представлена на видео.
 Средняя при этом должна оставаться свободной.
Средняя при этом должна оставаться свободной.Колосок на бок
Техника плетения колоска на бок на первый взгляд может показаться достаточно сложной. Подробная фото- и видео-инструкция поможет вам самостоятельно научиться выполнять такое плетение.
Расчешите шевелюру и разделите ее боковым пробором. Перекинь всю массу волос на одну сторону.
Разделите шевелюру горизонтальным пробором на две половины и начинайте плетение классического варианта колоска (подробную фото- и видео-инструкцию вы можете просмотреть выше). Когда коса достигнет линии ушей, продолжите плетение наискось (за ухо), притягиваю одну за другой пряди с левой и правой стороны.
Когда плетение дойдет до конца зоны роста волос, разделите волосы на три части и сплетите обычную косу. Зафиксируйте результат резинкой. Прическа колосок готова! Подробная фото- и видео-инструкция представлена ниже.
«Рыбий хвост»
Такой вариант выглядит очень красиво и изящно. Прическу можно выполнять и на тонких, и на густых локонах. Стилисты рекомендуют в процессе создания образа использовать тонкие пряди
, тогда результат получится очень нежным и аккуратным.
- Тщательно расчешите шевелюру. Разделите ее на две одинаковых половинки.
- С левую часть разделите на две половинки. Возьмите внешние локоны и перекиньте их через вторую половинку таким образом, чтобы они оказались по центру.
- Повторите шаг №2 справа.
- Продолжайте перекидывать пряди поочередно с обеих сторон. Помните, что от толщины взятых прядей зависит внешний вид прически. «Рыбий хвост», выполненный из мелких прядок выглядит очень элегантно и изящно.
- Продолжайте плетение до самых кончиков. Зафиксируйте результат резинкой. Прическа готова! Подробную технологию вы можете увидеть на фото и видео ниже.
 Возьмите внешние локоны и перекиньте их через вторую половинку таким образом, чтобы они оказались по центру.
Возьмите внешние локоны и перекиньте их через вторую половинку таким образом, чтобы они оказались по центру.Еще одна интересная модификация классического образа. Такой вариант прекрасно подходит для торжественных мероприятий. Двойная коса выглядит очень оригинально. С такой прической внимание окружающих обеспечено.
- Тщательно расчешите шевелюру. Выделите неширокую прядь в теменной зоне. Разделите ее на три одинаковые по ширине части.
- Начинайте работу в технике «рыбий хвост», поочередно подкладывая крайние пряди под центральную.
- На следующем этапе начинайте поочередно добавлять в основные пряди дополнительные локоны из основной массы волос.
- Затем из основной части вам необходимо выделить прядь, которая затем будет использоваться для создания второго «яруса». Для удобства зафиксируйте ее сверху заколкой.
- Сделайте подхват и сплетите косу под низ.
- Повторите шаги №4 и №5 с другой стороны. Продолжайте работу таким же образом до тех пор, пока не дойдете до зоны роста.
- Затем, когда уже все локоны заплетены, продолжайте работу, но без подхватов.
- Зафиксируйте косу резинкой.
- На следующем этапе необходимо заплести верхнюю косу. Для этого возьмите самую верхнюю свободную часть волос, разделите ее на три части и начинайте заплетать обычную косу. Затем постепенно выполняйте подхваты других свободных волос.
- Соедините две косы внизу резинкой. Прическа готова! Подробную технологию создания элегантного образа вы можете увидеть на видео ниже.
Коса колосок издавна пользуется популярностью среди женщин и девушек, это изысканный и одновременно простой способ привести шевелюру в порядок. Вариаций на эту тему много, от самой простой французской косы до изящных боковых и круговых причесок.
Вариаций на эту тему много, от самой простой французской косы до изящных боковых и круговых причесок.
Колосок – идеальный вариант для повседневного образа или для торжества. Научиться плести его довольно легко, можно делать эту прическу даже самостоятельно на своей голове.
Что необходимо для плетения
Очень оригинально смотрится колосок на мелированных или тонированных локонах.
Процедура плетения кос представляет собой чередование трех и более прядей, способы их объединения различны. Самое сложное в данном случае – удержать пряди, чтобы они не рассыпались, и правильно чередовать их.
При плетении не обойтись без вспомогательных средств, для создания красивого образа потребуются:
- Хорошая расческа, желательно деревянная, при расчесывании волосы не должны электризоваться;
- Заколки и зажимы, невидимки и резинки различных форм и размеров, они необходимы для фиксации отдельных прядей и самой косы;
- Средства для укладки и фиксации волос – гели, муссы, пенки, лаки;
- Ленты, банты и иные аксессуары для украшения косы.
Немаловажное значение имеет правильная подготовка волос. Опытные парикмахеры советуют предварительно тщательно вымыть шевелюру, желательно шампунем для придания волосам объема. Локоны от корней следует немного приподнять, для этого можно использовать фиксирующие средства.
Для создания ровной и аккуратной косички следует все пряди подбирать одинакового объема, желательно не толще мизинца. Коса из тонких прядей получается более тугой, лучше их отделять в направлении от уха к косичке.
Как сплести
Предварительно перед началом плетения необходимо волосы тщательно расчесать. Если шевелюра только вымыта, пряди следует хорошо просушить, чтобы они не были тяжелыми. На всю длину наносится пенка или мусс.
Это делать не обязательно, но обработанные пряди лучше ложатся и аккуратнее смотрятся. Слишком легкие волосы следует слегка начесать. Тонкие пряди можно слегка завить, чтобы прическа получилась более объемной.
Пошаговое руководство
Для плетения классического варианта колоска следует действовать следующим образом:
- На макушке отделяется ровная прядь волос, делится на три равные части. Условно их можно назвать 1, 2 и 3, если считать справа налево;
- Прядь 3 сверху заводится на вторую, первую накинуть на третью и подвести под вторую снизу;
- Вторая осталась с краю справа, к ней добавляется новая прядка с боку. Готовая прядь накидывается на соседнюю среднюю;
- Аналогичным образом захватывается новая прядь слева, соединяется с готовой и заводится на соседнюю посередине.
Захваты волос по бокам делать до тех пор, пока все локоны не будут задействованы в прическе. Оставшиеся волосы можно заплести как обычную косу, зафиксировать ее резинкой, бантом, заколкой.
Различные варианты плетения колоска
Существует немало различных методик создания колоска.
Для придания дополнительного объема и пышности, можно по окончании плетения расслабить и потянуть некоторые пряди.
Этот вариант смотрится очень оригинально, динамично и изящно, подходит для девочек и женщин. Волосы зачесываются назад, плетение начинается с любой стороны. Собирается прядь средней толщины, делится на три части. Начало плетения аналогично классическому способу.
Когда начинается забор дополнительных прядей, это делается только с одной стороны. Если колосок плетется справа, забор ведется с левой стороны. Маленькую прядь выделить из основной массы волос, перевернуть ее вокруг себя и соединить с крайней прядью.
Продолжать плетение, пока не задействованы все волосы. Можно делать заборы с двух сторон, при этом коса будет проходить вдоль уха с одной или другой стороны, закончить ее можно сзади у основания шеи, или продолжить дальше.
Кончик фиксируется резинкой или заколкой. Для того, чтобы сделать прическу более объемной и эффектной, из каждого звена можно слегка вытянуть прядки, а между ними можно вставить красивые шпильки.
Эта прическа выглядит очень шикарно, придавая образу неповторимый шарм и элегантность. Роскошный вариант подходит для торжественных случаев. Лучше всего она смотрится на густых волосах, если шевелюра негустая, от корней пряди лучше начесать.
Плетение производится следующим образом: локоны делятся на две равные части посередине пробором. Плетение начинается справа, выделяются три равные пряди, как для обычного колоска. Плетение ведется по спирали до тех пор, пока все пряди не будут задействованы.
Этот вариант выглядит более выразительно и эффектно, для его создания следует часть волос на макушке выделить, расчесать и разделить на три равные пряди. Плетение производится следующим образом: левая прядь заводится под центральную, правая – под левую и центральную.
С этого момента к каждой пряди прибавлять по равному количеству свободных волос, левая проходит под центральную, затем повторить то же самое с правой. Плетение продолжается по всей длине. Результат закрепить резинкой.
Пышный боковой колосок
Локоны разделить на неравные части косым пробором. Плетение начинать с одной или другой стороны, как обычный боковой колосок, с третьего плетения начинать захваты свободных прядей. Продолжить косу до мочки уха.
Затем с обратной стороны собрать волосы свободной рукой и скрутить их в тугой жгут, подвести его к косе. Оставшуюся часть косы можно закончить классически и воспроизвести . Результат должен быть с элементом небрежности.
Как сплести колосок самой себе
Самостоятельно сплести колосок самой себе довольно сложно без практики и сноровки. Чтобы добиться аккуратного результата, следует использовать средства для фиксации. Проще всего начинать плетение с бокового колоска, вообще всегда получается слегка растрепанным и объемным.
Проще делать косу из толстых прядей, она получается более объемной и слегка небрежной, некоторые огрехи в этом случае будут не так сильно заметны. Тонкие пряди позволяют создать изящный изгиб, на фоне которого самого плетения практически не видно.
Самостоятельно можно заплести двойной колосок. Для этого делается ровный пробор, и с двух сторон заплести по боковому колоску, сзади объединить обе косы в одну и доплести классическим способом.
Такая коса подходит практически всем, но немногие умеют правильно ее плести.
Для того, чтобы получить шикарный результат, можно воспользоваться советами и подсказками профессионалов:
- Для того, чтобы получилась изящная, аккуратная косичка, необходимо брать тоненькие прядки. Из толстых прядей получается прекрасная объемная коса;
- Если крупные пряди реже перекрещивать, получится слегка эпатажная прическа, которая в настоящее время считается очень модной;
- Чтобы избежать появление «петухов», каждую прядь перед перекрещиванием следует расчесывать и подтягивать;
- Проще заплетать слегка влажные волосы, можно намочить руки или обработать локоны муссом или пенкой.
Осваивая оригинальную и необычную прическу колосок, можно в полной мере проявлять свою фантазию, экспериментировать и добавлять интересные элементы. Это лучший вариант, где можно использовать любые , которые позволят сделать прическу еще более интересной и изысканной.
Красиво переплетённые косы, внешне напоминающие узор колоса, называются «Колоском». Его разновидностями считаются «Рыбий хвост», «Французская коса», а также множество двойных, тройных, воздушных косичек, корзинок и зигзагов.
Многие находятся в тупике, не понимая с чего начинать и как правильно плести колосок. Подробные пошаговые схемы помогут быстро научиться созданию эффектных причёсок.
Классический колосок является фундаментом для разнообразных косичек. Его создание занимает не более 3 минут. Освоив основную схему плетения, колосок можно видоизменять в различные причёски.
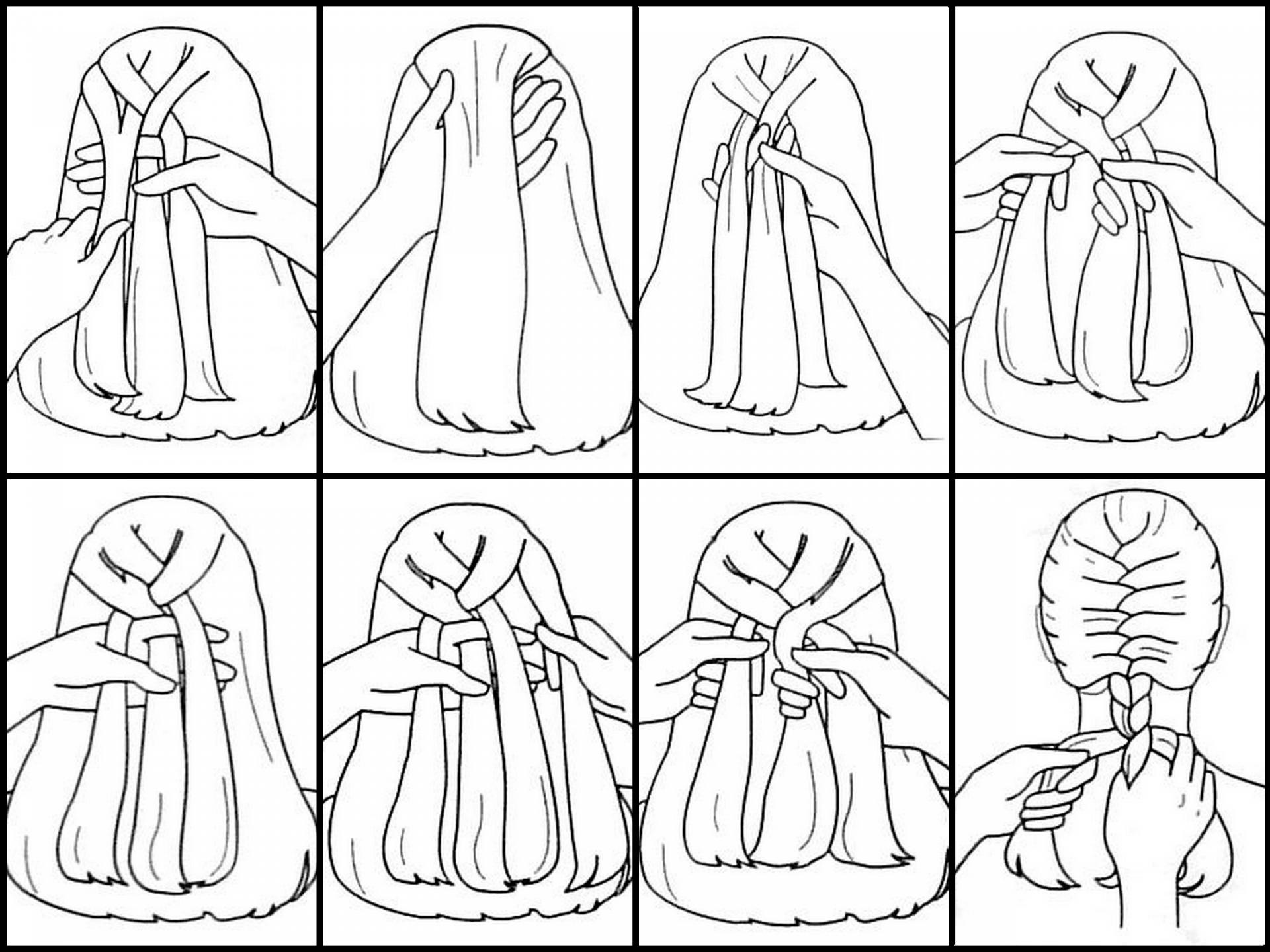
Принцип плетения несложный, но требует сноровки:
- Смочить волосы водой или увлажнить муссом, чтобы они стали послушнее и не путались в пальцах во время плетения.
- В области теменной части головы нужно отделить волос, толщиной в одну прядь. Новичкам рекомендуется собрать её в небольшой хвостик на макушке, закрепив резинкой. Это облегчит начало создания причёски и предотвратит её распад.
- С правой стороны отделяется вторая прядь такой же толщины и накладывается влево поверх собранной в хвост пряди.
- С левой стороны берётся третья прядь и накладывается на предыдущие переплетённые локоны.
- Затем необходимо отделить прядь от основной части волос, приложить к правому локону и продолжить плести.
- Далее взять локон с левой стороны головы и приложить к левой пряди.
- Поочерёдно брать одинаковые пучки волос слева и справа
- Плетение заканчивается обычной косой из 3 прядей.
- Колосок закрепляется красивой резинкой.
 Новичкам рекомендуется собрать её в небольшой хвостик на макушке, закрепив резинкой. Это облегчит начало создания причёски и предотвратит её распад.
Новичкам рекомендуется собрать её в небольшой хвостик на макушке, закрепив резинкой. Это облегчит начало создания причёски и предотвратит её распад.Как плести обычный колосок. Схема на фото.
Готовую косу можно завернуть внутрь и заколоть шпильками. Пряди аккуратно расправить, а косичку украсить вдоль небольшими заколками в виде цветов, побрызгать лаком.
Как заплести колосок самой себе?
Колосок можно заплести самой себе. Процедура займёт немного больше времени, так как пряди нужно отделять осторожно, чтобы они были ровными и аккуратными. Волосы перед началом работы нужно хорошо расчесать. Не рекомендуется постоянно смотреться в зеркало, так как это отвлекает внимание.
Инструкция:
- Необходимо осторожно отделить верхнюю часть волос и разделить её на 3 равных пучка.
- Затем начать плести обычную косу, накладывая прядь на прядь.
- После третьего накладывания локона, с боковой стороны отделяется небольшая часть волос и вкладывается в косичку.
- Последовательно накладывая одинаковые по толщине пряди, нужно доплести косу до основания шеи.
- Закрепить причёску резинкой.
Плетение кос на себе требует терпения, так как руки быстро затекают и колосок может получиться неровным. Поэтому новичкам для создания красивой косы нужно потренироваться.
Можно ли заплести колосок на коротких волосах?
Как плести колосок (схема для тупых покажет пошагово весь процесс плетения), так и создать другую прическу на коротких волосах невозможно без заколок – шпилек, невидимок, зажимов. Также поможет при распаде косичек тугое переплетение.
Также поможет при распаде косичек тугое переплетение.
Инструкция:
- Волосы смочить водой и расчесать.
- Из тонкой пряди начать плести обычную косичку. Чем тоньше пряди, тем лучше будут держаться локоны в причёске.
- Пройдя 6-7 рядов можно начинать брать локоны дополнительно, как в обычном колоске. Перекрещивать их нужно у корней, чтобы они не распадались. По ходу плетения начать прищеплять волос заколками.
- Затем закрепить аккуратно косичку резинкой и побрызгать лаком.
На короткий волос можно заплетать колосок, используя не весь волос, а только часть. Также создаётся причёска из двух или трёх косичек.
Пошаговые инструкции плетения разных вариантов колоска
Колоски очень разнообразны. Можно создавать любые красивые причёски, не ограничиваясь фантазией.
Колосок на бок
Колосок на бок смотрится необычно и эффектно.
Принцип плетения тот же, что и у классического колоска:
- Необходимо тщательно расчесать волос, направляя волосы назад, чтобы не было видно пробора.
- Затем аккуратно сбоку головы берётся прядь и делится на 3 равные части.
- Забор основной массы волос начинается после 3-4 перекрещивания локонов.
- Постепенно прикладывая волос с двух сторон к каждой пряди, нужно доплести косу до основания шеи и закрепить резинкой.
Косу на бок лучше плести из толстых прядей. Такой колосок смотрится оригинальней. При средней длине волос причёску нужно закреплять прищепкой или невидимкой. Коса на бок не подойдёт к короткой стрижке.
Наизнанку
Колосок наизнанку требует большей сноровки. В отличие от классического колоска, здесь пряди подкладываются снизу, а не сверху.
Как плести колосок (схема для тупых) наизнанку:
- Хорошо расчесать волос и сбрызнуть спреем, чтобы не путался.
- Отделить пучок ближе к чёлке.
- Разделить на 3 части.
- Начать плетение, протянув снизу левую прядь, между правой и средней.
- Повторить аналогичное действие с правой прядью.
- К левой пряди присоединить локон и протянуть его снизу вверх.
- К правой пряди также прикладывается волос.
- Продолжить плетение до основания шеи.
- Доплести обычной трёхрядной косой и закрепить резинкой.

Такая причёска имеет ярко выраженный узор, в отличие от обычного колоска. Коса выделяется на общем фоне и выглядит объёмной. Колосок наизнанку не очень подойдёт к коротким волосам, так как локоны будут постоянно выпадать.
2 вывернутых колоска
Подчеркнуть черты лица помогут 2 аккуратные косички. Необычно смотрятся вывернутые колоски по обеим сторонам головы.
Инструкция:
- Волос делится на 2 части. Одну часть собирается резинкой, чтобы не мешала.
- Далее нужно взять небольшой пучок и разделить на 3 одинаковых пряди.
- Каждый локон нужно подкладывать под другой, переплетая левый с центральным, затем правый с центральным.
- Пройдя 2-3 переплетения, можно начинать брать волос из основной массы.
- Закончив косу, нужно туго связать хвост резинкой, иначе из заколки волос может быстро выправиться.
- Вторую косичку сплести аналогично.
Косички по обе стороны головы подойдут как для длинного волоса, так и для средней и короткой стрижки. Колоски можно объединить в один хвост.
Вокруг головы
Колосок вокруг головы долго не распадается. Такая причёска подходит для садика и школы. Она плетётся на длинных и коротких волосах.
1 вариант для длинного волоса:
- Разделить волос на 4 равных части, сделав проборы от уха до уха и по центру головы от чёлки до основания шеи.
- Закрепить 3 части щипцами и начать плести классический колосок снизу вверх по часовой стрелке, аккуратно его закругляя. При плетении использовать 3 пряди, подбирая волосы с обеих сторон.
- Поднявшись до верхней закреплённой части волос, необходимо убрать щипцы, и продолжать плетение к чёлке.
- Когда волос закончится, нужно переходить на 3 часть, а затем на последнюю, четвёртую.
- Подобрав весь волос, оставшийся хвостик нужно доплести до конца, затем собрать его резинкой и спрятать, закрепив невидимкой.

2 вариант для короткого волоса:
- Расчесав волос сделать косой пробор от виска до макушки.
- Начать плести обычный колосок сверху от начала пробора, забирая пряди с обеих сторон.
- Пройдя весь круг и вернувшись вверх, до пробора, остаток волос нужно доплести до конца.
- Косичку уложить на пробор и закрепить невидимкой.
Колосок можно украсить красивыми шпильками и побрызгать лаком.
Змейка
Причёска «Змейка» состоит из одного классического колоска, который плетётся зигзагообразно.
При плетении берётся волос как с обеих сторон, так и с одной:
- Расчесав волосы назад нужно сделать косой пробор от виска в сторону макушки.
- Плетение начинается от виска, прядь разделяется на 3 небольшие одинаковые части.
- Волос при плетении колоска захватывается с обеих сторон.
- Достигнув уха, косичку нужно повернуть в другую сторону и плести, создавая треугольник с верхним колоском. Теперь дополнительные пряди подкладываются к основным только сверху, а нижние локоны остаются свободными.
- Закончив ряд, колосок снова нужно развернуть и продолжать плести, используя захват волос только сверху.
- Завершающий этап – плетение колоска до основания шеи и подбор оставшихся волос в одну простую трёхрядную косу.
Снизу волос собирается резинкой. Пряди можно аккуратно вытянуть, чтобы придать пышность «Змейке».
Колосок из хвоста
Колосок на хвосте называется – «Рыбий хвост». Сплести его поможет схема для тупых. Главный секрет этой причёски в том, что она создаётся из 2 прядей.
Инструкция:
- Необходимо собрать на макушке высокий хвост.
- Затем разделить его на 2 равные части.
- Осторожно отделить с внешней стороны правой пряди очень тонкий локон и положить его внутрь между двумя основными.
- Затем с левой пряди нужно взять тонкий локон и опять положить его между основными двумя. Важно аккуратно придерживать большими пальцами вплетаемые пряди.
- Внизу волос собирается резинкой.

Такая коса выглядит изящно, при этом она достаточно крепкая и быстро не распадается.
Коса в косе
Оригинально смотрится двойная коса. Она требует особой аккуратности при плетении.
Инструкция:
- Взять верхнюю часть волос и отделить прядь.
- Разделить прядь на 3 одинаковых локона.
- Начать плетение колоска наизнанку, подкладывая поочерёдно левую и правую пряди одну под другую.
- После трёх переплетений берётся волос из основной массы и добавляется в косу.
- От каждой большой пряди отделяется тонкая полоса волоса, которая остаётся свободной до конца плетения.
- Когда основная масса волос закончится, косу нужно доплести до конца, при этом также отделяя маленькие пряди.
- Закрепить колосок резинкой.
- Из заранее отделённых мелких локонов поверху плетётся тонкая коса наизнанку.
- Скрепить обе косы и немного вытянуть нижние пряди.
Для удобства, оставляемые мелкие пряди во время плетения нужно фиксировать специальной прищепкой.
Плетение 2 косичек
Косички с двух сторон всегда освежают лицо и особенно подходят молодым девушкам.
Их можно плести в разных техниках:
- простой колосок;
- вывернутая коса;
- рыбий хвост.
Инструкция:
- Разделить волос пополам, взяв одну из частей, а лишнее закрепить прищепкой или собрать в хвост.
- Начать плести от виска простой колосок, подхватывая полоски с обеих сторон. Если это косичка наизнанку, то подкладывать локоны нужно снизу, добавляя также пряди с обеих сторон. При технике «Рыбий хвост» пучок разделяется на 2 части и отделяемый с внешней стороны маленький локон закладывается между двумя основными.
- Когда дополнительных прядей для захвата не останется, плетение продолжается до конца в выбранной технике.
- Другая коса плетётся по той же схеме.

Не обязательно пробор должен быть строго посередине. Косой пробор подчеркнёт индивидуальность и поможет выделиться среди окружающих.
Рыбий хвост самой себе
Новичкам создать себе такую причёску будет непросто. Но существуют облегчённый способ – плетение на высоком хвосте.
Инструкция:
- Сделать хвост.
- Разделить его пополам и взять обе части в руки.
- Отделённый с внешней стороны левой пряди тонкий локон пропускается между двумя основными, и прикладывается к противоположному правому локону, тем самым создавая переплетение.
- Затем отделяется токая полоска с внешней стороны правой пряди и прикладывается к левой части. Чем тоньше локоны, тем изящнее выглядит колосок.
- Доплести косичку до конца и закрепить резинкой.
Освоив такой вид причёски, и научившись отделять одинаковые пряди, можно переходить к более сложным видам рыбьего хвоста. К ним относится плетение от чёлки или макушки с забором волоса, а также коса от затылочной части, где не нужно прихватывать дополнительные локоны.
Тройной рыбий хвост
Техника плетения этой модной причёски почти не отличается от двухрядного рыбьего хвоста. Она создаётся из трёх пучков.
Инструкция:
- С внешней стороны правой части берётся тонкая полоска и накладывается сверху на основной локон.
- Затем эта же полоска подводится под средней прядью и присоединяется к левой основной части.
- Далее тонкая прядь отделяется с внешней стороны левой части. Теперь она проводится под ней же и над средним локоном и прикладывается к правой стороне.
- Плетение продолжается до конца и закрепляется резинкой.
Получается, что средняя часть остаётся незадействованной. Она оказывается в кольце из тонких полосок, отделяемых с левой и правой основных частей.
Воздушный колосок
Для плетения косички нужно разделить весь волос на 3 части. При этом средняя прядь задействована не будет. Её нужно аккуратно придерживать во время всего плетения, регулируя натяжение косы.
Её нужно аккуратно придерживать во время всего плетения, регулируя натяжение косы.
Как плести колосок (схема для тупых в технике воздушного плетения):
- Отделив пучок на макушке, поделить его на 3 части.
- Правый локон перекидывается над центральным.
- Затем левый перекидывается над центральным.
- Далее захватывается полоска от основной массы и опять перекидывается через центральную прядь, которая постоянно должна находиться в руке.
- Действие повторяется с другой стороны.
- После того, как не останется прядей для захвата, коса плетётся в технике рыбий хвост. С внешней стороны прядей отделяются небольшие полоски и перекидываются через центральную часть, которая всё так же остаётся в руке.
- Закончив плетение, косичку нужно собрать резинкой и расправить звенья.
Такая коса подходит для длинных волос. Для придания причёске воздушности и объёма необходимо во время плетения выправлять звенья, немного вытягивая их и делая полукруглые воздушные узоры.
Волна
Волна создаёт колосок в виде ободка от одного уха к другому и плетётся на распущенный волос.
Инструкция:
- Аккуратно распределить волос по всей голове.
- Взять небольшой пучок с левой стороны от уха до чёлки и разделить на 3 локона.
- Начать плести классический колосок, захватывая дополнительные пряди с обеих сторон.
- Нижнюю прядь необходимо оставлять, а вместо неё добавлять в косичку дополнительные пряди.
- Такое плетение продолжается до середины головы.
- Затем точно также нужно сплести косу с правой стороны.
- Соединить обе косички и закрепить невидимкой.
Эта лёгкая обворожительная причёска подойдёт для длинного и среднего волоса.
Квадратный колосок на себе или двойной колосок
Причёска из крупных прядей выглядит элегантно и торжественно. Она заплетается на длинный или средний волос.
Инструкция:
- Взяв пучок волос в области темени нужно разделить его на 3 крупные части.
- Начать плести колосок наизнанку, подкладывая локоны под основные пряди.
- Постепенно захватывать волос с обеих сторон.
- Вплетая локоны в косу, нужно отделять с их внешней стороны крупную часть и закреплять прищепкой, чтобы не мешали.
- Доплести косу до конца и закрепить резинкой.
- Из оставшихся волос нужно создать вторую косу в той же технике.

В итоге должна получиться двойная коса – одна поверх другой. Чем крупнее оставленные пряди, тем ярче будет выражена квадратная форма косы.
Зигзаг
Для создания «Зигзага» нужна тонкая расчёска для проборов. Чем ровнее проборы, тем красивее будет смотреться причёска. Эта коса подобна колоску «Змейка». Разница в том, что при плетении простого колоска нужно захватывать дополнительный волос с обеих сторон.
Как плести колосок (схема для тупых в технике зигзаг):
- Сделать пробор слева от чёлки до правого виска и начать плести колосок, прихватывая волос с обеих сторон.
- Дойдя до правого уха, нужно заново сделать пробор наискосок к левому уху.
- Далее необходимо плести колосок в противоположную сторону, опять захватывая волос с двух сторон.
- Доведя косу до основания шеи, её завершают обычным плетением.
Закреплённый резинкой колосок можно украсить заколками и прищепками.
Диагональ
Коса по диагонали плетётся в разных техниках – классический колосок, колосок наизнанку, рыбий хвост. Начинать создавать такую причёску нужно с области виска.
Перед началом плетения необходимо расчесать волос и взяв прядь, разделить на 3 равные части. Плести косу нужно вниз по диагонали подбирая свободные волосы с обеих сторон и вплетая их в колосок. Если волосы средней длины, то желательно после окончания плетения подхватить волос с противоположной стороны невидимками.
Корзиночка
Красивая причёска, не выходящая из моды – это корзиночка. На первый взгляд она кажется сложной. Её можно делать на длинные и средние волосы.
Инструкция:
- Разделить волос на 2 части, сделав пробор от одного к другому уху и зачесав верхнюю часть волос вперёд.
- В центре на макушке нужно взять тонкую прядь и разделить её на 3 части.
- Начать плетение обычной косички по часовой стрелке, захватывая волос только справа.
- Когда волос закончится, косу собирают резинкой.
Хвостик нужно спрятать на голове, закрепив невидимками. Корзинку украшают заколками или прищепками.
Колосок с выпущенными прядями
Необычно смотрится коса с выпущенными прядями. Она построена на основе колоска наизнанку.
Инструкция:
- Сверху берётся прядь волос и делится на 3 части.
- Локоны переплетаются, подкладываясь под низ.
- Во время плетения основной волос прикладывается к косе.
- На каждой вплетённой пряди с внешней стороны отделяется локон и убирается в сторону. Действия выполняются поочерёдно справа и слева.
Дойдя до основания шеи, косу нужно доплести до конца, также выпуская локоны.
Крупный колосок
Крупный колосок плетётся как на длинных, так и на средних волосах. Он будет выглядеть тем красивее, чем гуще волос. Расчесав и расправив пряди, нужно отделить верхнюю часть волос и начать плетение вывернутой косы. Для этого нужно подкладывать пряди под низ, забирая поочерёдно локоны с 2 сторон.
Необходимо отделять крупные пряди, захватывая полностью волос от корней до косы.
Каждые 4 переплетения косы аккуратно расправляются. Так колосок будет выглядеть толстым и пышным.
Плетение с лентой
Украсить любую косичку можно красивой яркой лентой. Такую причёску можно сделать себе самостоятельно. Она плетётся в 4 пряди, одна из которых будет лентой. Начинать плести косу лучше с макушки.
Инструкция:
- Отделить верхнюю часть волос, разделив на 3 части.
- Закрепить невидимкой ленту под отделёнными волосами.
- Начать плести колосок наизнанку.
- Первую прядь положить под вторую и на ленту.
- Четвёртую прядь на первую и под ленту.
- Вторую прядь под четвёртую и на ленту.
- Первую на вторую, захватив волос с головы и провести под ленту.
- Четвёртую под первую, добавив волос слева и на ленту.
- Продолжать до основания шеи, пока не закончится свободный волос на голове.
- Затем доплести колосок до конца по тому же принципу.
Аккуратно закрепить ленту или завязать бантик.
Следуя подробным схемам даже самые неумелые смогут быстро научиться правильно заплетать колоски. Тонкие пряди сделают причёску аккуратной, а толстые локоны – объёмной и пушистой. Обладательницы средней и короткой стрижки могут не переживать за случайно выбившийся волос, так как небрежный стиль сейчас в моде.
Оформление статьи: Лозинский Олег
Видео о том, как плести колосок
Колосок. Техника плетения французской косы для новичков:
Косы — бессмертная классика, лучшее украшение женской головы. Это было актуально сотни лет назад, и остаётся модным сегодня.
Глядя на модных гламурных особ не отвратительной наружности, многие девушки задумываются о том, чтобы освоить искусство плетения колоска на собственной голове.
Это здорово: уметь за минуты превращать растрёпанные волосёнки в эталон женственности и красоты. Особенно здорово выглядят объёмные колоски с вытянутыми прядями.
Ажурная коса, спадающая с плеча — что ещё нужно, чтобы покорять сердца рыцарей, и вызывать приступы удушающей зависти у знакомых и подруг?
Только освоить плетение косы бывает непросто. Каждая девочка может заплести простую косу на другом человеке. А как это сделать на собственной голове?
Что, если попытки стать королевой красоты приводят к тому, что голова превращается в неаккуратное гнездо?
Не спешите стричь голову налысо, возьмите себя в руки. Есть способ научиться делать красоту своими руками.
В вашей власти научить руки плести настоящие шедевры:
- Во-первых, раз научились другие — сможете освоить искусство плетения кос и вы.
- Во-вторых, медведи в цирке не сразу научились управлять велосипедом.
- В-третьих, терпение и труд сделали из обезьяны человека, а из вас сделают мастера с чёрным поясом по плетению косичек любой сложности.
Вы станете настоящим гуру, способным за пару минут заплести колосок даже Дмитрию Нагиеву.
На себе, на другом, без расчёски, в невесомости или со связанными за спиной руками. Готовы приступить к обучению?
Схема плетения колоска на себе любимой — три шага к мастерству:
| № | Важные пункты | Подробности |
| 1 | Порядок действий | Приготовьте всё необходимое: расчёску, резинку или заколки. Слегка смочите голову, если на улице тепло или есть время высохнуть дома. Менингит не способствует развитию полезных навыков. Оденьтесь заранее, чтобы переодевание не сломало ваш хрупкий шедевр. Сядьте перед зеркалом в удобной позе |
| 2 | Полезные замечания | Учимся делать колосок. Зачешите локоны назад, возьмите верхнюю прядь рукой. Разделите на три части, переплетите между собой. Делайте медленно, чтобы понять, что получается. Затяните. Присоедините пряди с боков, старайтесь затянуть их туже, чтобы было ровно. Следите за толщиной: все пряди должны иметь примерно одинаковую толщину. Первые верхние пряди делайте толще, если хотите получить чёткую косу |
| 3 | Что ещё поможет рукожопу? | Если не умеете плести колосок на ком-то, бесполезно учиться на себе. Попросите подругу показать вам, как делается колосок, потренируйтесь на ней. Чем больше практики — тем лучше результат |
Как правильно заплести два колоска девочке
Плетение двух колосков — задача простая. Следуйте предложенной схеме, и будет вам счастье.
Схема плетения двух колосков:
- Расчёсываем локоны.
- Делаем вертикальный пробор, чётко разделяя голову на две одинаковые половины ровной линией.
- Закалываем половину волос, чтобы не мешалась. Это обязательно.
- Начинаем плетение от макушки на второй половине.
- Заплетаем косу, оставляя хвостик.
- Завязываем бантом или перетягиваем резинкой.
- Второй колосок делаем по образу и подобию первого.
Схема рыбьего хвоста для тупых
Рыбий хвост — разновидность косички, отличающаяся тонкими струйками, перетекающими друг в друга.
Для дам, с трудом освоивших обычный колосок, эта причёска покажется чем-то заоблачным.
Попробуем разобраться на примере фото-уроков. Ведь лучше один раз увидеть, чем сто раз услышать:
На рисунке, расположенном выше, указана схема выполнения рыбьего хвоста. Причёска уникальна тем, что даже в растрёпанном виде смотрится очень мило.
Она уместна на светском мероприятии, в офисе, на прогулке, на свидании и даже на субботнике. Универсальный вариант.
А здесь мы видим шедевр, выполненный наоборот:
Схема не отличается, только пряди заплетают не вперёд, а назад. Обратный метод. Но и обычные колоски с двух сторон будут выглядеть не менее прекрасно.
Прическа вокруг головы
Русская коса вокруг головы — что может быть красивее? С такой причёской ходили на свидание наши бабушки и прабабушки, очаровывая наших дедушек и прадедушек с первого взгляда.
Как сотворить такое на своей голове? Помните о медведях на велосипеде: им всё равно было сложнее. Вы освоите и этот способ.
Эволюционируем до мастеров: учимся делать круговой колосок:
- За такую работу берутся, предварительно освоив простой колосок. Москва не сразу строилась, и колосок не сразу плёлся.
- Потренируйтесь на других: мастерство заключается в практике. Откройте свой салон для подруг: приглашайте их в гости на мастер-классы по выдиранию волос. Вы сможете давать профессиональные уроки позднее.
- Для кругового плетения необходимо визуально перенести колосок в плоскость головы. Начните с затылка. Разделите голову на участки, чтобы локоны не спутывались.
- Пробуйте, пока не получится. Помните про обезьян: они долго трудились, прежде чем стать особями, которые учатся заплетать косички с помощью интернета.
Маленькая хитрость! Откроем секрет: наши бабушки вовсе не умели искусно заплетать колосок вокруг головы. Косы у девушек тех времён были толщиной с руку, до пояса и ниже.
Девушки заплетали обычную косу и укладывали её вокруг головы, чтобы досужие кавалеры не дёргали за неё.
Пошаговая инструкция косички, вывернутой наизнанку
Обратное плетение колоска: прекрасный способ создать плотную выделяющуюся косу.
Её размер будет зависеть от густоты волос. Но даже жидким локонам можно придать объём при помощи вытягивания прядей.
Обратимся к рисунку, где показана схема перестановки прядей:
Так делается колосок наоборот. Пряди не заходят на поверх косы, а поддеваются вниз. В остальном этот способ ничем не отличается от обычного колоска.
Вооружившись теоретическими знаниями, приступайте к практике. Не сдавайтесь при временных неудачах, пробуйте и всё получится.
Полезное видео
Как заплести колосок самой себе: схема плетения + видео
На чтение 4 мин. Просмотров 103k. Опубликовано Обновлено
Французская коса – безоговорочный тренд на все времена, вот только заплести ее на себе могут лишь единицы. Ситуация кардинально изменится после того, как вы прочтете нашу статью и узнаете, как заплести колосок самой себе.
Учимся сами плести колосок
Плетение сложных косичек самой себе считается делом очень нелегким. Но и опыт, согласитесь, играет огромную роль в делах красоты. Поэтому читайте наш мастер-класс и испытывайте его на себе.
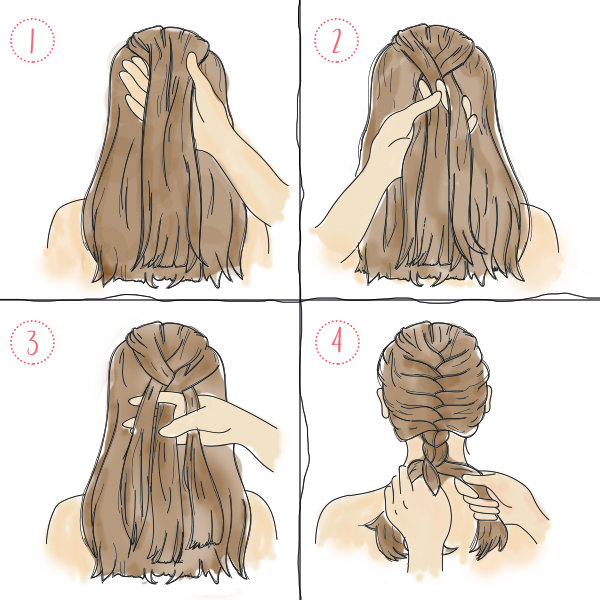
Шаг 1. Расчесываем волосы расческой.
Шаг 2. В верхней части головы отделяем прядь волос и делим ее на три равных части.
Шаг 3. Прядь №1 пропускаем между прядями №2 и №3.
Шаг 4. Прядь №3 кладем между прядью №1 и прядью №2.
Шаг 5. Прядь №2 располагаем между прядями №3 и №1. Тут же захватываем небольшую прядку справа и присоединяем ее к плетению.
Шаг 6. Крайнюю прядь кладем посредине и снова прибавляем пучок волос, но уже слева. Внимательно следите за тем, чтобы боковые прядки были одинаковой толщины, иначе прическа получится некрасивой.
Шаг 7. Повторяем два предыдущих пункта, продолжая плетение до основания шеи.
Шаг 8. Оставшиеся волосы заплетаем в обычную косу. Кончик завязываем резинкой.
Этот вариант французской косы считается самым простым. Если вы сумеете его осилить, то наверняка сможете покорить и прочие вариации на тему колоска.
[adsense]
Французский колосок наоборот
Схема плетения колоска наоборот самой себе может выглядеть сложной, но на практике все окажется очень легко.
- Тщательно расчесываем волосы, дабы во время плетения они не спутались.
- Отделяем небольшую прядь в верхней части головы и разделяем ее на три одинаковые секции.
- Захватываем первый локон (лучше слева) и заводим его вниз под две оставшиеся пряди.
- Третью прядь заводим под первую и вторую, то есть плетем косичку внутрь.
- Снова повторяем эти шаги, подбирая то справа, то слева дополнительные пряди (мелкие или крупные).
- Придерживаемся этого принципа до окончания длины волос, а кончик фиксируем резинкой.
- Желаете сделать колосок более объемным? Просто легонько растяните пряди руками.
Пышный колосок на бок
Колосок совсем не обязательно располагать посредине головы. С нашими подсказками вы легко заплетете его на бок и создадите романтичный образ.
- Расчесываем волосы расческой.
- Разделяем шевелюру косым вертикальным пробором.
- В той части, которая окажется больше, отделяем тонкую прядь и разделяем ее на три равные секции.
- Начинаем плести обычную трехрядную косу.
- На третьем плетении подключаем к колоску боковые пряди. Захватываем их то сверху, то снизу.
Шаг 6. Доходим до мочки уха и придерживаем кончик косички рукой.
Шаг 7. Волосы в противоположной части головы скручиваем в жгут свободной рукой. Двигаемся по направлению к косе.
Шаг 8. Соединяем обе части и доплетаем пряди в технике «рыбий хвост».
На выходе боковой колосок должен получиться растрепанным и объемным, так что об аккуратности плетения можете не волноваться.
Еще несколько модных вариантов плетение косы:
Плести французскую косу можно очень туго с использованием тонких прядок или же слегка небрежно, захватывая прядки потолще. Техника плетения одна и та же, но выглядеть оно будет совсем по-иному. Специалисты утверждают, что первый вариант идеально подходит юным девчонкам, тогда как второй лучше смотрится на взрослых.
А также, узнайте, как сделать красивые косички на длинные волосы своими руками.
Кому подходит коса по-французски?
Если раньше с колоском ходили только девочки в школе, то ныне его все чаще можно увидеть на взрослых. Эта укладка прекрасно смотрится на мелированных волосах, так как позволяет передать всю игру цвета. Среди невест особой популярностью пользуется колосок вокруг головы. Такая прическа выглядит просто шикарно, да и держится долго.
Теперь и вы знаете, как заплести колосок самой себе! Фантазируйте, сочетайте плетение с завитками, хвостами, жгутами и классической косой! Создавайте свои уникальные прически и не бойтесь экспериментов.
Как плести колосок пошаговая инструкция с фото
Свою первую косу я заплела лет в девять. До этого с моими волосами возились мама и бабушка. Да что там – даже дедушка мог сплести две простенькие косички. А у меня лично получалось лишь завязывать хвост. Просто волосы были длинные, очень густые и тяжелые – и справиться с ними своими силами ребенку было проблематично.
Тренировалась заплетаться я тогда, наверное, больше недели. И сердилась, и не получалось, и пряди выдирала. Но когда получилась толстая, ровная, аккуратно заплетенная коса – радости не было предела. И с того дня понеслось: каких только я причесок не выделывала! И водопад, и французские косички, и ажурные плетения по всей голове, и греческую косу, и венок, и сердце, и замысловатые узоры из 5-6 прядей.
Но удобнее и быстрее всего получался именно колосок. У этой прически множество плюсов:
— существует более сотни вариаций различных плетений – поэтому можно хоть каждый день ходить с новой укладкой,
— научиться колоскоплетению на самой себе проще простого – достаточно посмотреть видео инструкцию или схему заплетания,
— коса-колосок отлично подойдет для волос любого типа: тонкие и прямые волосы заплетаются вообще идеально, а вот с пышной шевелюрой придется повозиться, но результат того стоит.
Как плести колосок пошаговая инструкция с фото
Второе название колоска – «французская коса». Потому что именно во Франции первыми стали использовать такую технику плетения. На протяжении многих лет колосок то входил в моду, то незаслуженно отходил на второй план, уступая лидерство то каре, то кудрявым укладкам. Но в этом году коса-колосок снова в тренде.
Такая прическа прекрасно подходит и маленьким девочкам, и девушкам-красавицам, и деловым женщинам, и даже пенсионеркам. С прямым, классическим, угловым, ажурным колоском головка всегда будет выглядеть элегантно и эффектно. Эта прическа отлично впишется и в повседневный имидж, и идеально подойдет к любому праздничному наряду. И самое главное – за целый день такая прическа не расплетается, волосы ведут себя послушно. А после расплетания косы – на голове получаются прелестные волны.
Прежде, чем расплетать – колосок надо сделать. «Рецепт» простого колоска:
1) долго и качественно расчесаться, чтобы волос к волоску ложился.
2) разделить все волосы на две равные части.
3) определить точку начала колоска и взять там небольшую прядь волос, затем еще одну прядь с левой половины, а третью – с правой половины волос.
4) и начать плести как обычную косу: накладывая поочередно правую прядь на среднюю, а затем левую прядь на среднюю.
5) повторить накладывание прядей в основной косе раза 2-3, затем в основное плетение поочередно из правой и левой половин добавляем свободные пряди – то есть к нашей основной правой пряди мы добавляем свободную прядь и накладываем их на среднюю, затем тоже самое проделываем с левыми прядями.
6) важно помнить, что пряди должны быть равного объема, иначе может получиться неаккуратное плетение.
7) продолжаем плести до тех пор, пока все пряди не будут вплетены в косу, затем доплетаем до конца волос либо в технике классической косы, либо по методу плетения «рыбьего хвоста». И прическа готова.
Пошаговая инструкция для начинающих
Коса-колосок – это основа для всевозможных причесок. На базе колоска можно сделать и зигзаг, и венок вокруг головы, и двойной колосок, и русскую косу. Да хоть настоящее макраме из узорно заплетенных прядей и косичек. Но для новичков в колоскоплетении лучше всего начинать с самого простого. Для кого-то легче всего будет сделать прямой колосок. Для других может показаться удобнее, если делать плетение на правую или на левую сторону.
По сути, инструкция для начинающих ничем не отличается от рекомендаций, приведенных выше. Порядок действий аналогичный. Возможны лишь небольшие дополнения:
— каждую прядь, перед тем как вплетать, лучше всего расчесать несколько раз, чтобы волосы ложились аккуратно, и не было «гребешков».
— перед тем как начинать плести волосы, хорошо было бы потренироваться на тонких веревочках, чтобы отработать движения пальцев. Потому что если сразу практиковаться на волосах, то пряди будут путаться и это может доставить много неприятных ощущений модели, да и того, кто плетет, заставит изрядно поволноваться.
— хорошо бы посмотреть несколько разных уроков, изучить статьи-схемы, а уже потом, практически, выбрать для себя наиболее подходящий способ колоскоплетения.
— также можно на начальном этапе активно использовать резинки – с ними будет легко разделять прядки и манипулировать с волосами.
— неплохо было бы поставить зеркала, чтобы иметь возможность оценить плетение в процессе, и, если потребуется, внести сразу же коррективы, чтобы потом не переделывать все полностью.
Как сплести колосок самой себе
Чтобы делать колосок самой себе требуется приличная порция терпения и времени, но когда освоишься, и все движения будут доведены до автоматизма – на колоскоплетение будет уходить совсем немного времени.
На первых порах очень пригодятся заколки-крокодильчики – они помогут не путаться с прядями. Такая фиксация пригодится, если волосы длинные. Если волосы тонкие, то перед заплетанием, их стоит вымыть специальным шампунем, который придает объем. Для того чтобы коса-колосок смотрелась красиво, применяют также мусс или пенку для придания объема.
Пряди можно переплетать туго либо свободно. Для первого варианта нужны будут шпильки-резинки-заколки, лак для сильной или средней фиксации. Во втором случае можно обойтись без фиксации, и тогда укладка будет выглядеть слегка небрежно, но вместе с тем придавать определенный шарм. Лично у меня лучше всего получалось плетение от виска – косой колосок по всей длине волос.
Для самостоятельного заплетания лучше всего вначале освоить один вид – простой обычный колосок, а уже потом осваивать более сложные разновидности. А вообще не рекомендуется носить одну и ту же прическу каждый день – иначе волосы будут ломаться. Можно сделать так: сегодня, например, выплести красивую прямую косу, завтра – соорудить на голове оригинальный колосок из 4 прядей (тип плетения схожий, только пряди переплетают под две основные). Послезавтра сделать популярное плетение из двух колосков. В следующий раз правильное плетение из трех косичек, или крестообразное, или добавить ленты. Поле для экспериментов с образами – огромнейшее.
Колосок как плести видео
Создавать каждый день новую прическу – это приятно, уместно и стильно. Только к собственному стилю надо подходить очень ответственно. Если заниматься имиджем регулярно, то можно достигнуть определенных высот и даже перейти к следующему этапу: снимать на видео собственные мастер-классы.
Поделись с друзьями!
Следи за нами в соц. сетях!
Коса дракончик: как плести, варианты, фото
Если вы любите оригинальные прически с плетением, но традиционная косичка вам уже надоела, пришло время научиться делать дракончик. Техника выполнения его проста, поэтому вы сможете ее освоить самостоятельно, получив в результате стильный и запоминающийся образ.
История
Близкие родственники косы дракончик – колосок (французская коса) и африканское плетение. Именно от этих причесок он и ведет свою историю. Но если классический колосок обычно делается со спрятанными внутрь прядями, за счет чего он почти не возвышается над уровнем головы, то дракончик получается более объемным и выпуклым.
Прическа появилась в середине ХХ века, а пик ее популярности пришелся на конец 90-х – начало 2000-х годов. Сегодня этот вид косы снова становится все более актуальным.
Особенности
Преимущества дракончика:
- Подходит любой возрастной группе – причем не только женщинам, но и мужчинам. Дэвид Бэкхэм, Джастин Тимберлейк, а также многие RnB исполнители часто выбирают такую косичку.
- Хорошо вписывается во все стили – от эпатажного до строгого.
- Не распускается в течение дня. Это значит, что косичку можно делать на праздничные мероприятия, на которых вам нужно выглядеть элегантно на протяжении всего дня.
- Существует много вариантов выполнения косы.
Недостатки:
- Сильно стягивает шевелюру. Это может привести к головным болям.
- Не стоит заплетать слабые волосы, склонных к выпадению.
Для каких волос подходит?
Минимальной длиной, подходящей для создания дракончика, считается 10-15 см. Но если волосы короткие, воспользуйтесь канекалоном (синтетическими волосами). Прическа подходит как для прямых, так и для волнистых волос.
Можно делать не только на густых, но и на тонких и редких локонах. Если слегка распушить косу, вытаскивая из нее отдельные пряди, это придаст ей дополнительный визуальный объем.
Виды и процесс плетения дракончика
Существует много разновидностей косы, поэтому выбирайте вариант, который подходит вам больше всего.
Подготовка волос, аксессуары
Перед тем, как заплести косичку дракончик, нужно вымыть волосы, расчесать и нанести на них небольшое количество укладочных средств (пенки или мусса).
Для укладки, вам понадобится расческа-гребень для отделения прядей, резинка или заколка. Для дополнительной фиксации используется лак.
Классический
Освоив эту схему плетения, вы сможете создавать более сложные варианты дракончика.
- Отделите неширокую прядь надо лбом, разделите ее на три части.
- Возьмите левую прядь, перекиньте ее через среднюю, чтобы она оказалась крайней справа. Сделайте то же самое с правой прядью.
- Отделите тонкий локон в височной части слева и вплетите его в основную косу. Повторите это же с правой стороны.
- Вплетайте тонкие пряди с обеих сторон косы по всей длине волос. Далее плетется как обычная коса. В завершении зафиксируйте прическу резинкой.
Обратное (наоборот)
Техника плетения обратного дракончика отличается от классического варианта всего одной деталью. Первую левую прядь нужно проводить не НАД центральной прядью, а ПОД ней.
С двух сторон или два дракончика
- Разделите полосы прямым пробором на две равные части.
- Из рабочей зоны отделите три пряди и плетите классическим или обратным способом.
- Аналогичным образом сделайте вторую косу.
Главное требование к двусторонней косе в том, что при ее создании нужно брать пряди только одинаковой толщины. Левая и правая косы должны получиться симметричными.
Французский вариант
Второе название этой разновидности прически – дракончик набок. Начинайте плетение от виска и двигайтесь по диагонали в сторону противоположного уха. В результате вы получите косичку, пересекающее голову наискосок.
Вокруг головы
Коса делается вокруг головы, за счет чего получается эффект ободка или короны.
Отделите прядь надо лбом и разделите ее на три части указанным выше способом. Левую прядь проведите над центральной, добавляя волосы с правой стороны. Плетите косу вокруг головы, вплетая свободные пряди только с правой стороны.
Ажурный
Ажурный дракончик хорошо подходит для создания вечерних и свадебных причесок. Укладка характеризуется легкостью и воздушностью. Лучше всего коса выглядит на длинных волосах.
Схема плетения та же, что и в классическом варианте косы. Главное отличие заключается в том, что из готовой косички аккуратно вытаскиваются небольшие пряди одинакового размера. Для этого осторожно возьмите косу руками с двух сторон и равномерно тяните пальцами в противоположные стороны. В конце не забудьте сбрызнуть прическу лаком – иначе она может рассыпаться.
Как элемент прически (у лица, на висках и на макушке)
Дракончик может быть не только основной частью прически, но и входить в состав других. Например, вы можете заплести волосы на висках, челку или сделать окантовку вокруг пучка.
Коса с резинками
Для этого варианта дракончика вам понадобятся тонкие силиконовые резинки. Для светловолосых девушек подойдут прозрачные, для темноволосых – черные. Чем длиннее волосы, тем больше резинок вам потребуется.
- Расчешите волосы ото лба к затылку, отделите две неширокие пряди на висках и одну прядь на макушке. Все три пряди должны быть одинаковыми.
- Скрепите (не туго!) три рабочие пряди резинкой. Полученный хвостик перекрутите таким образом, чтобы он прошел в отверстие, образованное у основания резинки. В результате хвост должен «перекувыркнуться» вокруг резинки.
- Отделите две боковые пряди и соедините их с хвостиком, который будет играть роль центральной прядки. Снова закрепите полученный хвост резинкой.
- Продолжайте плетение до нужной длины. При этом следите за тем, чтобы каждый новый хвостик закрывал предыдущую резинку.
- В конце зафиксируйте косу красивой резинкой или заколкой.
Готовая коса выглядит очень эффектно, и подходит для торжественных событий – особенно если заплести ее на слегка подкрученные или гофрированные волосы и украсить эффектными аксессуарами.
Из хвостика с резинками
Укладка немного напоминает описанный выше дракончик с резинками.
- Сделайте небольшой хвостик на макушке. Временно заколите его парикмахерским зажимом, чтобы не мешал.
- Отделите две неширокие боковые пряди под первым хвостиком. Сделайте хвостик, аналогичный первому, и также закрепите его резинкой.
- Снимите зажим с верхнего хвостика. Разделите его на две равные части, между ними поместите волосы из нижнего хвостика и закрепите.
- Отделите две новые боковые пряди так, как было указано в пункте 2, сделайте еще один хвостик. Повторите указания, описанные в пункте 3. Заплетите таким способом все волосы.
- Старайтесь, чтобы каждый новый хвостик перекрывал собой резинку.
Полезные советы
- Чтобы сделать косу дракончик самой себе, воспользуйтесь трельяжем – он поможет вам хорошо видеть волосы сзади.
- Прическа сама по себе выглядит оригинально, но можно сделать ее внешний вид еще более интересным. Для этого используйте аксессуары – яркие заколки, ободки, искусственные и живые цветы, банты, повязки, шпильки, невидимки с бусинами, стразами и камушками. Отдельного внимания заслуживает коса, украшенная лентами.
- Можно внести в образ нотку романтичности, если слегка подкрутить щипцами хвостик.
- Дракончик является довольно тугим плетением. Делать его ежедневно не рекомендуется, так как это может привести к выпадению волос и головным болям.
- Если выполнять плетение на влажных волосах, после того, как вы распустите косу, получатся задорные волны.
- Если у вас жесткая шевелюра, с трудом поддающаяся укладке, воспользуйтесь специальными средствами – муссом или пенкой.
- Чтобы придать косе дополнительный объем, осторожно вытягивайте пряди из готовой прически, после чего сбрызните ее лаком.
- Чтобы создать аккуратную укладку, во время плетения берите пряди одинаковой толщины.
Мнение эксперта
Дракончик – красивый и эффектный вид плетения. Освоив его, вы сможете не только разнообразить арсенал своих ежедневных причесок, но и выглядеть стильно и привлекательно на торжественных мероприятиях.
Автор статьи: Bill Hedword
Тебе понравилось?…Поставь +1:
Как сделать красивую косичку быстро и легко. Как самой себе заплести косу – лучшие способы плетения. Обратная французская коса: легкое плетение
Современные причёски для детского сада и школы должны быть аккуратны и красивы. Как плести девочкам косы — это всегда актуальный вопрос для мам. Пошаговая инструкция, фото и видео помогут разобраться в этом вопросе. Причёски из кос для леди должны быть максимально безопасными, волосы не должны лезть ребёнку в глаза. Причёску хорошо закрепляем, особенно это касается малышей. Иначе мама к концу занятий может и не узнать дочку.
Начнём с причёсок на средние волосы для детского сада. Для этих причёсок делим волосы на дорожки, перекрещиваем их, оставляя хвостики внизу. Волосы держат цветные резиночки. Эти косички подходят на возраст 3-5 лет.
Для того чтобы каждый день делать девочкам красивые и модные школьные причёски, есть несколько советов:
- Малышка должна научиться сама хорошо расчёсывать волосы. Для этого понадобится качественная массажная щётка.
- Запаситесь подходящими к школьной одежде резинками и заколками разных размеров и форм. Обратите внимание: чем гуще и длиннее волосы, тем крепче должны быть заколки.
- Нужны специальные детские шампуни и бальзамы. С длинными волосами хорошо справляется спрей для облегчения расчёсывания.
Обратите внимание, что косичка (хвост и узел) ни в коем случае не должны быть слишком тугие! Это нарушает кровообращение головы и может плохо отразиться на самочувствии ребёнка. Причёски в школу должны быть аккуратными и не должны стягивать кожу головы.
Выбивающиеся пряди на висках лучше закрепить невидимками. Сейчас в продаже есть много ободков-резинок и тонких, и широких. Такая незаметная резинка сможет удержать все непослушные пряди возле головы.
Ещё одна причёска из кос для школы. Есть очень много вариантов, но мы возьмём основной вариант — называется он французская коса или колосок. Начинаем плести колосок с верхней точки головы. Просто берём прядь волос и разделяем её на 3 части. Перекрещиваем пряди так, как будто вы делаете обычную косу. Затем к переплетённым прядям добавляем боковые. Важное замечание: чем тоньше пряди, тем изящнее получится колосок. В конце заканчиваем простой косой и заколкой или резинкой для волос. Если вы посмотрите на косу, то поймёте почему причёска называется колосок. Очень похоже на колос.
Ещё одна простая, но очень оригинальная причёска из кос для девочек — закрученный хвост. На возраст 6-10 лет. Сделать её своими руками можно, потратив всего 5 минут. А потребуются пульверизатор с водой, расчёска и две резинки для волос. Немного смочим волосы, чтобы они лучше ложились. Расчешите шевелюру и соберите в хвост, закрепив его резинкой.
Разделите волосы на три части. Они должны получиться одинаковыми.Сначала одну прядь разделите пополам, скрутите два жгутика и соедините их.Проделайте то же самое с оставшимися прядями. Соедините все пряди вместе, закрепив резинкой. Косичка закрутится сама собой. Получается красивая причёска, которую можно при желании украсить бантом.
Французская косичка
Способ номер 1
Ещё одна причёска на основе французской косы. Для неё нам понадобится резинка, расчёска, аксессуары для украшения, пульверизатор для воды. Слегка смочить волосы. Расчешите и разделите пробором. Отделите небольшую прядь волос у левого виска.
Затем разделите её на 3 части и сплести полуколосок. Это значит, что захватывать прядки нужно только снизу. Доплести косу по всей голове малышки, закрепить резинкой и украсить лентой.
Способ номер 2
Заберите волосы со лба и разделите их на 3 прядки. Один раз переплетите,как обычно. Одним пальцем правой руки захватите тонкую прядку справа от правой прядки косы. Присоедините ее к правой пряди и переплетите со средней прядью, как обычно. Таким же образом, одним пальцем левой руки заберите тонкую прядку с левой стороны, присоедините к левой прядке косы и переплетите со средней прядкой, как обычно.
Продолжайте плести косу, захватывая тонкие прядки с обеих сторон попеременно. Когда свободно ниспадающие волосы закончатся, продолжайте плести косу, как обычно до самых кончиков, закрепите её резинкой. Когда научитесь плести «Французскую косу», то сможете плести косички в любой комбинации. Следите за тем, чтобы пряди были примерно одинаковой толщины.
Другие варианты причёсок из кос
- Бохо стиль. Богемные косички в основном подразумевают свободный стиль, то есть они отлично подходят для повседневного образа, гармонично дополняя его и придавая ему изюминку. Одной из самых популярных кос бохо является плетение вокруг головы, только главное, не спутать бохо с колоском. Косички бохо украсят причёску любой девочки. Весьма модным будет заплетать косички на распущенных прядях, то есть получится эдакий свободный образ, в котором волосы могут быть уложены хаотичным порядком, а вот сама коса будет лежать безупречно.
- Коса пучок. Она стала популярной не так уж и давно, представляет она собой соединение воедино двух модных укладок. Коса пучок – это, прежде всего стиль и элегантность. Для того, чтоб создать безупречный образ, нужно сперва нанести на волосы специальный мусс, делающий их гладкими и послушными.
- Коса колосок. Заплетается «колосок» посередине, по-другому он также называется «дракончик». Плетётся он наоборот для того, чтобы получился объёмный эффект. Как вариант, можно делать плетение с лоскутками ткани, что будет очень оригинально и нетривиально.
- Коса вокруг головы. Такая коса была популярной уже давно, во многом благодаря своему романтичному виду. Для того, чтобы сделать подобную причёску, необходимы расчёска, несколько шпилек, а также запас терпения. Делается она двумя способами: два хвоста, которые соединены в области позади ушей; колосок, в него вплетены боковые пряди волос. Оба варианта выглядят очень стильно и придутся по душе школьницам.
Мы с вами узнали как плести косы девочкам и какие способы существуют. В конце хотелось бы показать, как сделать причёску для торжественных случаев. Украшаем красивыми бантами и аксессуарами.
Причёска для девочки на праздник или семейное торжество.
Похожие записи:
Коса – символ женственности во все времена
С давних времен коса отождествлялась с женской красотой. Косы часто украшают классические, современные прически. Популярность их постоянна, не зависит от возраста женщины, цвета ее волос. Основные способы заплетения косичек известны давно. Придумать новое уже невозможно, но современные парикмахеры умудряются преподносить сюрпризы обладательницам длинных волос.
Можно абсолютно точно сказать, что заплетенные волосы сегодня являются предметом украшения.
От древности до наших дней
До наших времен дошли наскальные рисунки с изображениями причудливых форм на головах. В прически традиционно вплетались украшения, которые указывали на принадлежность к племенам. У Самсона из древних преданий вся сила была в волосах. Но хитрая спутница Далила обрезала семь его прядей и лишила могущественной силы.
В эпоху Возрождения прически перестали скрывать под головными уборами. Женщины начали подчеркивать свою красоту и индивидуальность, заплетая волосы. В моде были прически с множеством переплетенных мелких косичек.
Интересно, что именно в этот период стали отбеливать волосы, используя для этого настой ромашки и сушку волос на солнце. Украшениями служили яркие ленты, жгуты, богатые дамы вплетали жемчужные нити. На полотнах известных художников можно увидеть модниц той эпохи.
Русские красавицы традиционно носили косы. Первые упоминания относятся еще к домонгольскому периоду. На Руси одним из признаков семейного положения девушки было количеству кос. Незамужние девушки заплетали одну косу, после того как выходили замуж делили ее на две.
После петровских реформ у знатных женщин такая форма укладки не пользовалась популярностью. Они отдавали предпочтение европейской моде и припудренным парикам. А люди попроще продолжали заплетать косы. Они любили их из-за простоты и удобства.
Мир косичек
Сегодня косички переживают второе рождение. Одинокая тоненькая переплетенная прядь будет к лицу совсем юным созданиям. Леди постарше такой вариант не подойдет. Загадочность образа подчеркнет легкая растрепанность, замысловатое переплетение прядей. Мелкие косички могут асимметрично присутствовать в небольшом количестве. Они придадут легкий шарм, подчеркнут моложавость обладательницы.
Длина волос, как и цвет, может быть разная. Чем длиннее волосы, тем большие возможности предоставляются парикмахеру для фантазии и творчества. Но если захотелось заплести косу, а волосы не очень длинные, то это вполне возможно.
Прекрасно выглядят маленькие косички на коротких волосах. Они могут украсить челку или быть в виде короны или венка. Пользуются популярностью френч-водопады, заплетенные вокруг всей головы или с какой-нибудь одной стороны. Для создания этой прически подойдут волосы средней длины.
Модный бренд – необычное заплетание косичек.
Классическими вариантами стали: французские, датские, греческие, «рыбий хвост» и африканские косички. Для создания элегантного и женственного образа молодой особы сделайте французский или греческий вариант. Если предстоит активный отдых, остановите свой выбор на форме «рыбий хвост». Он также подойдет для любительниц стиля casual. Если вы обладательница кудрявых и прямых волос, попробуйте прическу в греческом стиле.
Подчеркнуть достоинства, скрыть недостатки
Лица бывают самых разных форм: круглые, овальные, длинные, квадратные. Большое значение имеет расположение и цвет глаз, размер носа, высота лба. Рот может быть в форме сердечка, большим и маленьким. Правильно подобранная прическа должна украшать, подчеркивать красивые черты и корректировать недостатки.
- Если обладательница узкого лица будет отращивать волосы и заплетать их в косу, то только акцентирует внимание на вытянутой форме. Более подходящей будет не высокая прическа, волосы, мягко падающие вокруг щек, которые визуально округлят форму лица. Корни лучше не открывать, а волосы не поднимать вверх. Остановитесь на волнистой укладке и средней длине волос с небольшим пробором. Освежить такую прическу можно, заплетая несколько маленьких косичек типа френч-водопад.
- Девушкам с треугольной формой необходимо придать объем нижней части лица. Это можно сделать с помощью удлиненной асимметричной челки. Дополнит прическу классическая коса, начинающаяся от затылочной части головы.
- Женщинам с прямоугольной формой лица пойдет широкая и длинная коса. Короткий волос не сможет скорректировать форму лица. Французская коса или колосок от шеи визуально уменьшит такое лицо.
Дамам с овальным лицом пойдут все виды кос. Этот тип внешности наиболее универсален для любых причесок. Если обладательница еще имеет и длинные волосы, то экспериментировать можно бесконечно. Все виды причесок с использованием кос будут смотреться естественно и элегантно. - Круглое лицо необходимо искусственно удлинить, поэтому косичку надо начинать плести в области макушки. Обычно заплетают на всю длину волос, до самих кончиков.
Густота шевелюры также имеет значение при выборе прически. Ведь у каждой женщины она индивидуальна. Это зависит от цвета, расы, здоровья. Больше всех волос у блондинок, а меньше у рыжеволосых красавиц. Главным условием остается все же правильный уход за ними и хорошее состояние организма. Добавить объема можно с помощью французской косы. С ней можно экспериментировать как угодно, располагая в разных частях головы.
Плетение косичек – простор для фантазии
Многие народы используют разные виды плетения. Но в основе каждого лежит переплетение определенного количества прядей. Классическая коса состоит из трех прядей, на которые делится весь объем волос.
- Для прически «рыбий хвост» используется две части из общего объема волос и техника плетения отличается от классического способа.
- Французский вариант предполагает равномерное вплетение прядей, которые распределены по всему объему головы.
- Датская коса, популярная последнее время, является противоположным вариантом французской прически или «колоска». Она плетется снизу, а пряди подкладываются одна под другую.
- Африканский стиль плетения завоевал популярность у молодежи. Он наиболее затратный по времени. Очень часто прическу украшают разными цветовыми оттенками.
- Швейцарская коса отличается простотой исполнения и эффектным видом. Техника не отличается от классического варианта плетения. Если вы захотите плести косу из плотно скрученных прядей, то в результате получится швейцарский вариант прически.
Правильная техника для заплетения волос
Современная мода на прически очень демократична. Трудно утверждать, будто существуют какие-то законы плетения кос. Точно, можно сказать, что правильно плести косички из чистых, тщательно расчесанных волос. При этом человек, делающий прическу должен быть аккуратен, чтобы не травмировать кожу головы.
Для этого необходима определенная сноровка и ловкость рук, а опыт придет со временем. Если есть еще и модель, желающая обновить прическу, то все обязательно получится. Главное условие для получения качественного результата это одинаковое натяжение по всей длине.
Чистый волос тщательно расчешите. Затем поделите на три равные части и начните переплетать. Попеременно меняя положение левой и правой прядей, перемещая их в центр, у вас будет получаться коса.
В зависимости от того, сколько косичек вы хотите получить, разделите волосы пробором на части, каждую из которых затем разделите на три и заплетите в косу. Если все волосы разделить на три одинаковых по толщине пряди, то получится одна коса. Отделив прядь волос у виска, можно заплести односторонний «колосок», который будет красиво обрамлять лицо. При этом можно закрепить конец косички противоположного виска, а можно доплести ее до конца.
Во время плетения косы, слишком слабая плотность приведет к тому, что прическа быстро потеряет форму. А чересчур сильное натяжение волос может вызвать дискомфорт.
Полет фантазии при создании причесок и немного терпения сделают вас неповторимой, подчеркнут красоту и вашу индивидуальность. Коса позволит выглядеть женственно в праздник и будни.
Переходим к действиям – фото и инструкции
Лучше всего, конечно, посмотреть видео выше о том, как плести косички, но если нет такой возможности, можно сделать несколько прекрасных вариантов по следующим инструкциям, причем самой себе и в домашних условиях (нажмите на картинку, чтобы открыть на полный экран).
Время для современных модниц очень ценно, поэтому прическу хочется сделать как можно быстрее и эффектнее! В этой статье собраны 17 самые красивые, а главное быстрые прически на каждый день для каждой длинны — средней, длинной и каре.
Легкая прическа самой себе за 5 минут из переплетенных жгутов пошагово:
Вам понадобится:
- 3 резинки в цвет ваших волос (используйте мелке силиконовые для косичек)
- заколка для украшения (пожеланию)
- лак для фиксации по желанию
Выделите верх волос, свяжите резинкой и протяните резинку сквозь собранные волосы. Выделите еще две пряди по бокам и свяжите их ниже под первым оборотом. Сделайте этими прядями несколько оборотов. Свяжите волос в один хвост ниже всех оборотов. Украсьте любимой заколкой ил оставьте как есть. Эффектно, легко быстро, самая простая каждодневная прическа невероятной красоты.
Красивая прическа с косами за 5 минут самой себе:
Как сделать легкую прическу самому себе на средние волосы с косами:
- шпильки/невидимки
- силиконовые резинки
- зажимы
Выделите переднюю часть волос (от уха до уха) закрепите зажимами на лбу. Заплетите косу с лева на право, растяните сплетенные пряди закрепите косу за ухом. Выделенные волосы спереди сплетите в такую же косу и закрепите шпильками ли невидимками под нижней косой, ее плетите с права на лева.
Легкий красивый пучек на длинные волосы за 10 минут самой себе:
Для того чтобы сделать пучек самой себе вам потребуется:
- невидимки
- расческа для начеса
- резинка
Завяжите высокий хвост, взбрызните лаком и начешите пряди. Соберите волосы в ту форму пучка, которая вам по душе и закрепите невидимками.
Очень красивая прическа на длинный волос, которую легко сделать самой себе дома:
Для простой прически на длинный волос в домашних условиях вам нужно взять:
- шпильки/невидимки
- силиконовую резинку
- средства фиксации по вашему усмотрению
Разделите волос на 3 части параллельно друг другу (для средней части выделите чуть больше волос). Заплетите колосок обратку, растяните пряди косы и заплетите хвостик внутрь косы. Возьмите пряди, которые вы оставили с боку и хаотично вплетайте в основную косу. Фиксируйте их невидимками внутри основной косы.
Красивый хвост для длинных не очень густых волос быстрая прическа для выполнения самой себе дома:
Для того чтобы сделать легкий красивый хвост на каждый день вы должны подготовить:
- силиконовые резинки в цвет своих волос
Завяжите хвост сзади или сбоку, сделайте несколько оборотов резинкой через волос. Завяжите ниже еще одну резинку и проделайте тоже самое, так до самого окончания волос.
Бант из волос за 10 минут самой себе дома пошагово:
Как сделать бант из волос, что нужно для этого дома:
- шпильки/невидимки
- резинка
Завяжите высокий хвост, протяните его под резинкой, растяните бант такого объема, как желаете оберните остаток кончиков вокруг резинки зафиксируйте невидимками.
Гулька — поэтапно сделать самой себе за 3 минуты для волос средней длинны:
Что потребуется для гульки в домашних условиях:
- шпильки
- невидимки
Соберите волосы в хвост, скрутите их в жгут, оберните в бублик кончик волос закрепите под гулькой шпилькой и невидимкам.
Простое плетение на каждый день для длинных волос поэтапная инструкция по фото:
Чтобы сделать самой себе плетение из волос цепочкой вам нужны:
- силиконовые резинки
Завяжите хвост. Возьмите 2 пряди по бокам и оберните об среднюю часть хвоста, завяжите резинкой.Повторите такие действия до окончания длинны волос и вытяните все пряди в конце. Прическа выглядит супер, вы потратите на нее 10 минут, но окружение будет в замешательстве, как вы самс ебе умудрились создать такое плетение.
Трехсторонний колосок в домашних условиях самой себе пошагово фото. Прическа на длинный волос:
Чтобы быстро и просто заплести себе колосок по трем сторонам подготовьте:
Заплетите обычный обратный колосок, но оставьте по бокам от висков по одной небольшой пряди (очень тоненькой) завяжите косу резинкой. После возьмите две пряди и закрутите в спираль, зафиксируйте спираль по длине косы и растяните.
Легкая прическа — пучек из прядей поэтапно фото:
Вам необходимо подготовить для прически:
- шпильки/невидимки
- резинка
- локон
- зажим
Соберите волосы в хвост и накрутите локоны крупно. Зафиксируйте одну прядь и начните оборачивать вокруг нее пряди оставшиеся в хвосте. Ваша задача россыпью связать все пряди хвоста упаковывая во внутрь и зафиксировать их невидимками.
Прическа на длинные волосы красиво легко быстро:
Как сделать красивую и легкую прическу на длинный волос самой себе очень быстро:
- нужны резинки для косичек
Выделите верхнюю часть волос шапочкой и завяжите в хвост, протяните локон хвоста через резинку, завяжите ниже еще одну прядь добавляя еще пряди и повторите все как на фото. Оставшуюся длину спрячте под укладку.
Легкая прическа розочка из косы на длинные и средней длины волосы самой себе пошагово фото:
Как сделать розочку из волос, что нужно для этого:
- резинка
- шпильки/невидимки
Заплетите косичку из трех прядей по длине волос до конца волоса, соберите косичку бубликом и зафиксируйте ее на начале плетения. Кончики накрутите локоном.
Легкая домашняя укладка на каре самой себе пошагово фото:
Вам потребуется:
- шпильки/невидимки
- резинки
Соберите верхнюю часть волоса и завяжите в хвост, прокрутите через резинку так сделайте до тех пор, пока не получится собрать гульку. Нижние пряди поднимайте к гульке и фиксируйте невидимками, оставляя гульку открытой. Часть в которой собраны все пряди украсьте бантом или заколкой.
Красивая легкая прическа на каждый день для кучерявых волос средней длинны:
Как легко уложить вьющийся волос в укладку а 5 минут самой себе поэтапно:
- шпильки/невидимки
Выберите область на верху головы для начала плетения косы, заплетите обычный колосок до средины головы, закрутите и подберите оставшиеся волосы к косе в пучек. Наслаждайтесь красивейшей прической и делайте ее легко по фото инструкции.
Домашняя быстрая укладка на волос средней длины как сделать самой себе:
Как быстро сделать прическу за 5 минут самой себе:
- резинки
- украшение
Возьмите 2 пряди волос и свяжите их сзади, возьмите 2 следующие пряди и завяжите над предыдущей резинкой, так несколько раз, всю композицию плетения окончите украшением.
Как собрать волосы в красивое плетение дома смой себе быстро и красиво:
Чтобы быстро заплести самой себе прическу из двух кос возьмите:
Разделите волосы на 2 части (верхнюю и нижнюю) заплетите косы на одну строну как для водопада и сплетите между собой в спираль.
Как сделать красивую прическу самой себе инструкция в фото пошагово дома:
Вам нужны:
- резинка
- невидимки
Завяжите хвост исключая передние части, их оставьте для плетения. Берите прядь, которую оставляли и прядь с хвоста, переплетайте в косу, по окончанию заплетите все по окружности.
Легкие прически видео:
О том, как заплести себе красивую прическу без посторонней помощи, уже написано много статей и снято множество видео-роликов.
Так почему бы не объединить самые популярные из них, чтобы под рукой всегда были лучшие способы создать новый образ в домашних условиях?
Самой универсальной прической во все времена всегда считалась косичка.
Наверняка многие из вас умеют плести самый ее простой вариант из трех прядок, а в детстве мама заплетала на ваших локонах и французский вариант косы.
Процесс довольно простой, однако справиться с ним на своих собственных волосах куда сложнее, чем плести косичку на чужой голове.
Однако, всего несколько простых шагов помогут вам решить эту задачу, и вы сможете хоть каждый день радовать себя новыми прическами с косичками и делать их самой себе.
Прежде чем перейти непосредственно к инструкциям и видео-урокам, следует приготовить необходимые инструменты, которые понадобятся вам для плетения: зеркало удобной величины, чтобы вы могли хорошо следить за процессом, спрей или вода для сбрызгивания локонов, разные расчески, аксессуары для закрепления: невидимки, резинки, заколки, шпильки.
Для фиксации результата, возможно, вам понадобится лак. В случае, если прическу вы делаете для повседневной носки, то ее не обязательно фиксировать таким образом.
Плетем жгут
Самая простая косичка, с которой можно начать тренироваться заплетать косы самой себе – это жгут.
Плетется очень быстро, выглядит оригинально:
- Для начала хорошенько расчешите все прядки и соберите их в хвост на макушке. Сделайте его потуже;
- Поделите локоны на две секции и начинайте по отдельности скручивать каждую в тугой жгут. Необходимо скручивать их в одном направлении – по часовой стрелке или против – выбор за вами;
- Теперь нужно переплести их между собой по спирали;
- Закрепите результат резинкой для волос.
Французская косичка своими руками
Французская косичка всегда выглядит очень романтично и красиво. Научиться делать ее самой не сложно. Достаточно попрактиковаться, и у вас в кармане новая прическа.
Просто повторяйте простые шаги:
- Прядки расческой отводим назад;
- Выделите одну небольшую прядь на макушке и поделите на три равные секции;
- Начинайте плести классическую косу, но при этом добавляйте с каждым звеном по небольшой прядке с боков. Необходимо задействовать все волосы;
- Как только все волосы будут вплетены, продолжаем плести обыкновенную косу до кончиков волос, либо можно оставить распущенные волосы;
- Результат закрепите резинкой и сбрызните лаком для волос.
Французская косичка будет смотреться очень красиво с дополнительными аксессуарами: в голову можно вставить цветы или само плетение дополнить атласной лентой.
Односторонний ободок
Если вы хотите создать образ романтичной, милой девушки, то вам наверняка понравится такая прическа:
- Отделите локоны надо лбом от остальных волос так, чтобы рабочая область была не сильно толстой;
- Начинаем плести тонкую косичку от уха. Постепенно добавляем прядки к этой косе, как и для французской косички. Нельзя брать волосы с макушки;
- Необходимо довести до следующего уха и там закрепить незаметным образом и прикрыть распущенными волосами;
- При желании можно собрать остальные локоны в нетугой пучок, тем самым создав небольшую небрежность и еще более романтический образ.
Микс из косичек
Такая прическа будет прекрасно смотреться на длинных волосах.
Подойдет как для работы, так и для романтического свидания.
- Отделяем прядки боковым пробором;
- Заплетите с одной стороны французскую косичку, вплетая распущенные волосы по очереди;
- Как только дошли до уха, закрепите косичку резинкой;
- С другой стороны собираем оставшиеся волосы и делаем жгут;
- Соединяем с косичкой и закрепляем резинкой;
- Оставшиеся локоны можно заплести в классическую косу или «рыбий хвост».
Эта прическа может показаться вам сложной, однако делается она довольно просто. Вам просто нужно несколько раз попрактиковаться, и вы обязательно добьетесь идеального результата.
«Рыбий хвост», ободок и интересный микс
Поверьте, даже с такой сложной задачей вы сможете справиться самостоятельно и заплести «рыбий хвост» самой себе:
- Поделите расчесанные волосы на две секции поровну;
- Возьмите тонкую прядку с левой стороны и перебросьте ее вправо;
- Повторите действие с прядкой, но уже с правой стороны. Обратите внимание, что чем тоньше прядки, тем лучше будет смотреться такой вариант косы;
- Таким же образом продолжайте плетение до получения косички нужной длины;
- Закрепите результат резинкой.
Это идеальный вариант для прически на работу. Однако она занимает немного больше времени, по сравнению с плетением французской косички.
Но если у вас уходит немного времени на сборы по утрам, то вполне можно заняться созданием и такой прически.
Можно сделать довольно простую прическу с ободком по всей голове.
Для этого необходимо, чтобы у вас были волосы ниже лопаток:
- Расчешите прядки и поделите пополам ровным пробором. Хотя здесь можно пофантазировать и сделать какой-нибудь оригинальный пробор: волной или зигзагом, рваный;
- Теперь нужно заплести себе классические косички с обеих сторон на одном уровне, начиная от уха;
- Возьмите одну косу и перекиньте ее надо лбом к противоположному уху;
- Хорошо закрепите невидимками не только конец, но и всю косу по длине, чтобы ободок оставался на месте;
- Тоже самое проделайте с другой косичкой, только на затылочной части;
- Места крепления можно украсить цветами или красивыми заколками.
Это идеальный вариант для школьницы или для любительниц активного образа жизни. Делается довольно просто, все волосы принимают участие, а значит, они не будут мешать вам в течение всего дня.
Однако почему бы не совместить два несложных плетения в одно, чтобы создать неповторимый образ?
Простая классическая косичка легко совмещается с французской.
Вот простые шаги, как это сделать:
- Делаем боковой пробор и хорошо расчесываем прядки;
- Берем прядку у самого лица и плетем одностороннюю французскую косичку. Это значит, что захватывать дополнительные прядки необходимо не с двух сторон, а только с одной. Необходимо захватить не все волосы, а лишь небольшую часть;
- Возле уха начинаем плести обычную трехпрядную косичку;
- Закрепляем ее резинкой;
- Остальные локоны собираем на тот же бок, где у нас тонкая «полуфранцузская» коса и плетем вместе с ней обычную классическую косичку;
- Чтобы сделать плетение ажурным и объемным, аккуратно вытягиваем петли, но не до конца.
Как видите, заплести косички самой себе совсем несложно. Достаточно лишь попрактиковаться и следовать каждому шагу в инструкции.
Подробные видео-уроки помогут вам освоить любую технику плетения: от самой простой до сложнейшей. Поэтому не бойтесь экспериментировать.
Наберитесь терпения, возьмите в руки расческу и начинайте делать себе самые красивые прически самостоятельно.
Коса — это довольно универсальная и практичная прическа, любимая многими девушками. Первые упоминания о таком виде плетения волос можно найти в Библии, где рассказывается о том, что Самсон носил 7 кос, в которых находилась вся его сила. Также косички носили в Древнем Египте и Греции.
На Руси этой прическе уделялось особое внимание. Если у девушки была длинная толстая коса, то она считалась сильной и здоровой. К тому же существовала традиция, согласно которой незамужние девушки носили одну косу. Ее украшали всевозможными лентами и цветами. А вступая в замужнюю жизнь, им заплетали две косы, которые сверху соединялись в корону.
Опрятная коса делается по нескольким несложным правилам, следуя которым легко поразить всех.
- Идеально расчесанные волосы — первый залог идеальной косы.
- Чтобы прическа была аккуратной пряди должны быть одинаковыми.
- Отделять пряди нужно равномерно, а также обязательно оттягивать вниз, чтобы они получились ровными по всей длине.
- Нужно следить за натяжением прядей, иначе коса получится местами слабая, а местами слишком туго заплетенная.
- Перед плетением заранее необходимо приготовить расчески, зажимы, резинки, средства укладки, а также различные аксессуары для украшения будущей косы.
На сегодня существует множество видов косичек, которые отличаются не только по своему внешнему виду, но и по сложности плетения. Причем это могут быть косички, с которыми можно ходить несколько недель, а также повседневные прически.
Разновидностей плетения сегодня достаточно много, чтобы выбрать ту или иную технику для каждого случая. Самые простые варианты подойдут для ежедневного стиля, работы, путешествия. А вот более сложные техники станут настоящей изюминкой образа на торжественных мероприятиях или свидании.
Итак, какие же есть виды косичек и в чем их особенности?
Дреды
Дреды — это целенаправленно спутанные пряди, которые могут быть разной толщины и длины. Само плетение дредов может быть разным.
Первый метод заключается в разделении волос на квадратики и начесывании их против роста в направлении к корням. А затем выбившиеся волоски вплетаются в пряди при помощи крючка. Полученные прядки натираются специальным воском, чтобы они хорошо держались.
Второй способ — это простое скручивание прядей и закрепление их на конце резинкой. Результат также закрепляется воском.
И третий — это перманентные дреды, которые делаются только у мастеров. Для этого используются химические средства, которые обеспечивают «жизнь» плетения в течение нескольких месяцев.
Эти оригинальные косы-трубочки по стилю подойдут далеко не каждой. Они не вписываются в строгий или деловой стиль, не подходят для торжественных случаев. Но зато могут радовать в повседневном образе, да и ухода особенного не требуют.
С помощью специального «вяжева» оплетаются небольшие прядки собственных волос, таким образом становясь более массивными, с эффектом начесанности.
Когда волосы поделены на пряди, они закрепляются у корней нитями определенного цвета. И затем этой же нитью начинается плотное без просветов обматывание волос. После того как несколько сантиметров пряди заплетены, можно сменить цвет нитки. Концы волос можно дополнить яркими бусинками.
Зизи
Это уже готовые тонкие косички быстрого плетения. Они могут быть прямыми, гофрированными, волнистыми и спиральными. Преимуществом такого плетения является то, что оно подходит тонким негустым волосам.
Толщина зизи достигает всего 3мм, они крепятся к натуральным заплетенным волосам. Поэтому на их плетение уходит2-4 часа в зависимости от скорости работы мастера и изначальной длины волос. К тому же цвет зизи может быть любым, так как их плетение предполагает перекрытие исходного цвета волос.
Керли
Керли имеет вид крупных тугих локонов, срок службы которых достигает 2-3 месяцев при правильном уходе. По цвету такие вплетаемые локоны не могут слишком отличаться от натурального оттенка собственных волос, иначе это будет выглядеть неестественно.
Техника плетения зависит от изначальной длины волос. Длинноволосым понадобится блейд, то есть плетение тонких кос, плотно прилегающих к голове. В них вплетаются готовые локоны керли. Точечное вплетение керли необходимо для коротких волос, которые не требуют, чтобы их прятали.
Особенностью этих косичек является то, что на концах они не заплетены, а имеют как бы хвостик, от куда и происходит их название. Свободные хвостики могут быть прямыми или накрученными.
«Пони» крепятся на свои волосы, заплетенные в мелкие тугие косички. Длина таких прядок может быть любой, к тому же это отличный способ удлинить свои волосы.
На создание классических афрокосичек уйдет много времени, а также понадобится специальный материал канекалон, который будет вплетаться в волосы.
Афрокосички — это тонкие косички по всей длине волос в количестве от 150 до 200 штук. Они могут иметь два вида хвостиков — прямой или завитый. Для этого волосы делятся на мелкие прядки, после чего начинают плестись тонкие равномерные тугие косички. При этом центральные пряди натягиваются на себя, а боковые к левому и правому уху соответственно.
Косички на каждый день
В настоящее время косички очень актуальны, к тому же они уже долгое время не выходят из моды. Существует много видов плетения, благодаря которым можно создавать разные образы. Для многих плетение косичек — довольно увлекательное и легкое занятие. Здесь все зависит от практики, которая поможет справляться даже со сложными плетениями быстро и аккуратно.
Французская коса
Именно эту технику плетения любят модные подиумы, часто используют знаменитости, да и простые девушки. Коса в французском стиле выглядит очень нежно, делается легко и не испортит даже деловой образ. У нее есть несколько видов плетения, которые можно подбирать под случай.
Сперва нужно задать начало косы, это три равных по толщине локона, как для самых стандартных техник. Далее плетение также напоминает простую косу, но вместо того, чтобы плести локоны строго по длине, они идут по кругу головы. При сплетении каждого нового ряда, прядки подбираются от соседних локонов, чтобы получился эффект венка, или боковой косички. В зависимости от желания, можно плести ее туго или слабо, от этого будет зависеть более строгий или романтический вид прически в итоге. По завершении плетения нужно закрепить косичку резинкой
Если вы все еще не знакомы с техникой плетения французской косы, а фото-уроки кажутся вам непонятными, смотрите мастер-класс на видео ниже. Наглядный пример поможет разобраться!
Колосок
Второе название этого плетения – «рыбий хвост». Вначале волосы зачесываются назад или сделать пробор. Затем, по желанию делается хвост, либо же, можно сразу приступать к созданию прически. По бокам, чуть выше висков, отделяются по одному локону. Один локон накидывается на другой, и по мере продвижения вниз по бокам также захватываются отдельные прядки. Держать такую косу нужно в натяжку, и добавляемые прядки должны быть одинаковой толщины, чтобы прическа получилась аккуратной.
Данную технику знает каждая славянская девушка, но не каждая может с ней справиться из-за нехватки практики. На самом деле, плести обычную русскую косу проще простого. Три одинаковых по толщине локона сплетаются между собой. Каждый раз во внутрь плетения идет то один, то второй боковой локон. А прядь, которая оказывается в этом случае центральной, просто придерживается пальцами. При частой практике заплести себе русские косички можно менее чем за минуту не зависимо от длины стрижки.
Коса «Дракончик»
Классическая техника плетения «дракончика» напоминает французскую косу, но начинаться она должна прямо ото лба. В другой похожей технике, косичка начинает походить на настоящий хвост дракона. Она становится более рельефной, плетется туго и также может называться «перевернутая коса». В этом случае крайние пряди нужно не накладывать сверху, а, наоборот, вплетать под среднюю. Так следует проделать со всей длиной переходя вниз по голове. Натяжение прядок можно немного уменьшить, аккуратно вытянув их. Тогда объем косы увеличится еще больше.
Коса «Водопад»
Коса «водопад» плетется горизонтально от самого начала пробора у лба. Итак, плетение начинается с деления у висков начальных трех локонов одинаковой толщины. Прядки перекидываются друг через друга, при этом одна из них отпускается и остается с общей массой волос, а вместо нее берется новая верхняя прядь. Плести волосы нужно до противоположной височной части головы, после чего закрепить косу невидимкой под волосами. Таким образом, распущенные волосы и будут создавать тот самый водопад. Их можно слегка накрутить, чтобы придать прическе романтичности.
Здесь используется 4 пряди, которые плетутся между собой поочередно. Для удобства создания этой косы в одну руку берется по две пряди, и начинают скрещиваться между собой первая со второй, третья с четвертой. В завершение скрещиваются между собой пряди, которые находятся в середине. Таким образом плетение идет по всей длине волос, после чего можно закрепить концы красивой резинкой или прочной заколкой.
Этот вариант косы достаточно сложный, но смотрится она очень интересно и оригинально. Лучше всего ее плести с самого верха волос. Итак, поделив затылочную часть волос на 5 частей, они будет перекрещиваться таким образом — первая прядь идет второй, но под низ третьей, и с другой стороны повторить тоже самое — пятая под четвертую и сверху образовавшейся третьей. Остальные ряды плетения делаются по аналогии.
Коса жгут
Такая косичка достаточно оригинальная, а ее техника кардинально отличается от плетения стандартной косы, что, впрочем, не делает ее более сложной в исполнении. Базой служит конский хвост, который делится на две равные части волос. Они обе туго скручиваются в жгуты. После этого полученные жгутики закрепляются резинками. Затем можно приступать к перекручиванию частей, но уже в противоположную сторону. Резинки с двух частей снимаются и прическа закрепляется.
Коса с лентой
Ленту можно вплести в любой вид косы. Для этого необходимо под одной прядью сделать маленький хвостик, к которому и прикрепляется лента, которая будет подходить к образу.
Есть еще один вариант, который состоит из четырех частей, одна из которых будет состоять из ленты, прикрепленной к центральной пряди. Плетение будет таким же, как при косе из четырех частей, но лента должна идти строго по центру, как бы закрепляя прядки.
Девушки с такой удобной длиной волос легко смогут заплести себе все виды доступных кос. Для них подойдут привычные виды — колосок, жгут, французская.
Удобная и не сложная прическа, которая уберет волосы от лица и откроет шею — греческая коса. Плетется она как обычная, но только идет внизу по кругу головы, закрепляется невидимками.
Как научиться на себе плести колосок
Колосок можно заплести на волосы от 12 см. Этот вид косы одинаково хорошо смотрится как на волосах средней длины, так и на очень длинных. В процессе плетения можно использовать дополнительные ленты или банты: это выглядит красиво и нарядно. Учиться плести колосок легче всего на другом человеке. Когда вы освоите эту технику, сможете без труда заплетать косу и себе.
Вначале хорошо расчешите волосы и нанесите на них немного фиксирующего средства. Например, это может быть воск. Таким образом, пряди не будут распадаться и выпадать из рук. Для удобства возьмите в помощь расческу с длинной ручкой, которой можно разделять волосы на пряди.
Начинайте плетение с теменной зоны или макушечной. Вначале разделите волосы на три пряди. Если у вас есть челка, отчешите ее на лоб и заколите, чтобы не мешала. Начните плетение, сделав 2-3 звенья косы. После этого продолжайте заплетать колосок, но к каждой боковой пряди делайте подпряд то с левой, то с правой стороны. Внимательно следите за прядями, чтобы не перепутать их.
Кладите в центр прядь за прядью, а также соединяйте их ровно, чтобы колосок получался красивым и рельефным. Следите за тем, чтобы волосы в косе были одинаковыми по толщине. От этого также зависит красота плетения. Конечно, из косы не должны выбиваться волоски, не должно быть петухов и торчащих прядей. Для этого старайтесь заплетать колосок туго и подтягивать пряди, если они ослабевают.
Продолжайте плетение до тех пор, пока позволяет длина волос. В самом конце остается небольшой кончик волос: его можно не заплетать. Чтобы колосок не расплелся, зафиксируйте его резинкой, заколкой, крабиком или шпильками.
Если вы хотите сделать колосок объемным, ослабьте косу. Делается это так: одной рукой придерживайте косу снизу, а второй рукой попеременно ослабляйте прядки. Учтите, что вытягивать прядки нужно симметрично и на одинаковое расстояние. Кое-где выбившиеся волосы зафиксируйте невидимками. В завершение можно нанести на прическу лак для волос.
Колосок не обязательно располагать прямо по центру. Его можно заплести набок, по диагонали и даже волной, начиная от макушки и опускаясь ниже. Очень красиво на волосах выглядят два опускающихся колоска. Конечно, для такой прически у вас должны быть достаточно густые волосы. Если вы хотите вплести дополнительную ленту, соедините ее с одной прядью и следите, чтобы она не перепуталась с остальными. Тогда коса получится очень красивой.
Home | СинфлоресценцииStapleton (1997) и другие интерпретировали колоск (то есть маленький колос) как представляющий основной тип соцветия в семействе злаковых; эта интерпретация здесь принята. Это означает, что скопление колосков становится сложной структурой, которую мы называем синфлоресценцией.Синфлоресценции злаков традиционно описывают как шипы, кисточки или метелки, или как некоторые разновидности одной из этих тем. Однако следует отметить, что в дополнение к сложному характеру синфлоресценции травы они созревают по определенному образцу, так что травы, включая бамбук, не имеют настоящих колосьев, кистей или метелок, за исключением самих колосков, которые являются прицветниковые шипы. У бамбука также существует дополнительная проблема интерпретации псевдоспиклетов, которые содержат бутоносные (гемморальные) прицветники, которые обычно продолжают развиваться дополнительные порядки псевдоспиклетов, но в стиле цимоза.Чтобы сделать вывод о гомологии синфлоресценции бамбука, необходимо более глубокое понимание эволюции бамбука, полученное из других источников данных, поэтому символы в этом разделе сосредоточены в первую очередь на структурах, которые различают колоски и псевдоколоски. С-1. Один или несколько гемматериальных прицветников, образующих собственно колоск: 0 = отсутствует; 1 = присутствует, бутоны развиваются впоследствии или нет. С-2. Отступающий прицветник в основании оси, несущей колос или собственно колос: 0 = отсутствует; 1 = присутствует. С-3. Морфология сдвигающегося прицветника: 0 = чешуевидный или присутствует в виде рубца или ободка, длиной не более нескольких мм, лезвие отсутствует; 1 = хорошо развиты как с ножнами, так и с лезвием (часто видоизменяется). С-4. Профилл у основания оси, несущей колос или собственно колоск: 0 = отсутствует; 1 = присутствует. С-5. Профиллы: 0 = целиком; 1 = по крайней мере, несколько глубоких трещин, которые можно разделить вдоль на две половины. С-6. Один или несколько покровных прицветников, связанных с пучками колосков или собственно колосков: 0 = отсутствует; 1 = присутствует. Продолжительность жизни и корневища |
Генетическая модификация расположения колосков пшеницы увеличивает количество зерен без значительного влияния на массу зерна
Alaux M, Rogers J, Letellier T, Flores R, Alfama F, Pommier C, Mohellibi N, Durand S, Kimmel E, Michotey C, Guerche C, Loaec M, Lainé M, Steinbach D, Choulet F, Rimbert H, Leroy P, Guilhot N, Salse J, Feuillet C, Paux E, Eversole K, Adam-Blondon AF, Quesneville H, Consortium IWGS (2018) Linking эталонная последовательность генома мягкой пшеницы Международного консорциума по секвенированию генома пшеницы в соответствии с генетическими и феномическими данными пшеницы.Genome Biol 19: 111
Статья
CAS
PubMed
PubMed Central
Google Scholar
Alonso MP, Mirabella NE, Panelo JS, Cendoya MG, Pontaroli AC (2018) Селекция по высокому индексу плодовитости колоса увеличивает генетический прогресс в урожайности зерна и стабильности мягкой пшеницы. Euphytica 214: 112
Статья
CAS
Google Scholar
Alvaro F, Isidro J, Villegas D, del Moral LFG, Royo C (2008) Старые и современные сорта твердой пшеницы из Италии и Испании различаются по основным компонентам колоса.Полевые культуры Res 106: 86–93
Статья
Google Scholar
Aoki N, Whitfeld P, Hoeren F, Scofield G, Newell K, Patrick J, Offler C, Clarke B, Rahman S, Furbank RT (2002) Три гена транспортера сахарозы экспрессируются в развивающемся зерне гексаплоидной пшеницы. Растение Мол Биол 50: 453–462
Артикул
CAS
PubMed
Google Scholar
Ассенг С., Эверт Ф., Мартр П., Роттер Р.П., Лобелл Д.Б., Каммарано Д., Кимбалл Б.А., Оттман М.Дж., Уолл Г.У., Уайт Д.В., Рейнольдс М.П., Олдермен П.Д., Прасад ПВВ, Аггарвал П.К., Анотай Дж., Бассо B, Бирнат С., Чаллинор А.Дж., Де Санктис Дж., Долтра Дж., Феререс Е., Гарсия-Вайл М., Гайлер С., Хугенбум Дж., Хант Л.А., Изаурральде Р.С., Джаблун М., Джонс С.Д., Керсебаум К.С., Келер А.К., Мюллер К., Кумар С.Н., Нендель К., О’Лири Дж., Олесен Дж. Э., Палосуо Т., Призак Е., Резаи Е. Е., Руане А.С., Семенов М.А., Щербак I, Стокл С., Стратонович П., Стрек Т, Супит I, Тао Ф, Торберн П.Дж., Ваха К., Ван Э., Уоллах Д., Вольф И., Чжао З., Чжу Й. (2015) Повышение температуры снижает мировое производство пшеницы.Nat Clim Change 5: 143–147
Статья
Google Scholar
Austin RB, Bingham J, Blackwell RD, Evans LT, Ford MA, Morgan CL, Taylor M (1980) Генетические улучшения урожайности озимой пшеницы с 1900 года и связанные с ними физиологические изменения. J Agric Sci 94: 675–689
Статья
Google Scholar
Beales J, Turner A, GriYths S, Snape JW, Laurie DA (2007) Регулятор псевдоответа неправильно экспрессируется в нечувствительном к фотопериоду мутанте Ppd-D1a пшеницы ( Triticum aestivum L.). Theor Appl Genet 115: 721–733
Статья
CAS
PubMed
Google Scholar
Blade SF, Baker RJ (1991) Реакция веса ядра на изменения поглотителя источника в яровой пшенице. Crop Sci 31: 1117–1120
Статья
Google Scholar
Boden SA, Cavanagh C, Cullis BR, Ramm K, Greenwood J, Finnegan EJ, Trevaskis B, Swain SM (2015) Ppd-1 — ключевой регулятор архитектуры соцветий и развития парных колосков у пшеницы.Nat Plants 1: 1–16
Статья
CAS
Google Scholar
Bonnet OT (1967) Соцветия кукурузы, пшеницы, ржи, ячменя и овса: их зарождение и развитие, том 721. Университет Иллинойса, Бюллетень экспериментальной станции Колледжа сельского хозяйства, Шампейн
Google Scholar
Borras L, Slafer GA, Otegui ME (2004) Реакция сухой массы семян на манипуляции с источником – поглотителем пшеницы, кукурузы и сои: количественная переоценка.Полевые культуры Res 86: 131–146
Статья
Google Scholar
Borrás L, Slafer GA, Otegui MaE (2004) Реакция сухой массы семян на манипуляции с источником – поглотителем пшеницы, кукурузы и сои: количественная переоценка. Полевые культуры Res 86: 131–146
Статья
Google Scholar
Chapman JA, Mascher M, Buluc A, Barry K, Georganas E, Session A, Strnadova V, Jenkins J, Sehgal S, Oliker L, Schmutz J, Yelick KA, Scholz U, Waugh R, Poland JA, Muehlbauer GJ, Stein N, Rokhsar DS (2015) Подход с использованием полногеномного дробовика для сборки и закрепления гексаплоидного генома мягкой пшеницы.Genome Biol 16:26
Статья
PubMed
PubMed Central
Google Scholar
Кокрам Дж., Джонс Х., Ли Ф. Дж., О’Салливан Д., Пауэлл В., Лори Д. А., Гренландия А. Дж. (2007) Контроль времени цветения злаков умеренного пояса: гены, одомашнивание и устойчивая продуктивность. J Exp Bot 58: 1231–1244
Статья
CAS
PubMed
Google Scholar
Craufurd PQ, Wheeler TR (2009) Изменение климата и время цветения однолетних культур.J Exp Bot 60: 2529–2539
Статья
CAS
PubMed
Google Scholar
Cruz-Aguado JA, Reyes F, Rodes R, Perez I., Dorado M (1999) Влияние соотношения источника и стока на разделение сухого вещества и 14 C-фотоассимилятов в пшенице во время налива зерна. Ann Bot 83: 655–665
Статья
Google Scholar
Debernardi JM, Lin H, Chuck G, Faris JD, Dubcovsky J (2017) microRNA172 играет решающую роль в морфогенезе колосьев пшеницы и обмолоченности зерна.Разработка 144: 1966–1975
Статья
CAS
PubMed
PubMed Central
Google Scholar
Dixon LE, Greenwood JR, Bencivenga S, Zhang P, Cockram J, Mellers G, Ramm K, Cavanagh C, Swain SM, Boden SA (2018) TEOSINTE BRANCHED1 регулирует архитектуру и развитие соцветий у мягкой пшеницы ( Triticum aestivum ). Растительная клетка 30: 563–581
Статья
CAS
PubMed
PubMed Central
Google Scholar
Добровольская О., Понт С, Сибут Р., Мартинек П., Бадаева Е., Мюрат Ф., Шоссон А., Ватанабе Н., Прат Е., Готье Н., Готье В., Понсе С., Орлов Ю. Л., Красников А. А., Бергес Н., Салина E, Laikova L, Salse J (2015) FRIZZY PANICLE запускает лишние колоски в мягкой пшенице.Plant Physiol 167: 189–199
Статья
CAS
PubMed
Google Scholar
Ellwood ER, Temple SA, Primack RB, Bradley NL, Davis CC (2013) Рекордно раннее цветение в восточной части США. PLoS ONE 8: e53788
Артикул
CAS
PubMed
PubMed Central
Google Scholar
Фаричелли М.Е., Валарик М., Дубковский Дж. (2010) Контроль времени цветения и развития колосьев у зерновых: раннеспелость per se Eps-1 область у пшеницы, риса и Brachypodium .Funct Integr Genom 10: 293–306
Статья
CAS
Google Scholar
Ферранте А., Савин Р., Слафер Г.А. (2012) Различия в физиологии урожайности между современными, хорошо адаптированными сортами твердой пшеницы, выращиваемыми в контрастных условиях. Полевые культуры Res 136: 52–64
Статья
Google Scholar
Ferrante A, Cartelle J, Savin R, Slafer GA (2017) Определение урожайности, взаимодействие между основными компонентами и стабильность урожайности традиционной и современной пшеницы в широком диапазоне сред.Полевые культуры Res 203: 114–127
Статья
Google Scholar
Фишер Р.А. (1985) Количество зерен в посевах пшеницы и влияние солнечного излучения и температуры. J Agric Sci 105: 447–461
Статья
Google Scholar
Фишер Р.А. (2007) Понимание физиологических основ потенциальной урожайности пшеницы. J Agric Sci 145: 99–113
Статья
Google Scholar
Flintham J, Börner A, Worland A, Gale M (1997) Оптимизация урожайности зерна пшеницы: эффекты карликовых генов Rht (нечувствительные к гиббереллину).J Agric Sci 128: 11–25
Статья
Google Scholar
Фоулкс М.Дж., Слафер Г.А., Дэвис В.Дж., Берри П.М., Сильвестр-Брэдли Р., Мартр П., Кальдерини Д.Ф., Гриффитс С., Рейнольдс М.П. (2011) Повышение потенциала урожайности пшеницы. III. Оптимизация разделения зерна при сохранении устойчивости к полеганию. J Exp Bot 62: 469–486
Статья
CAS
PubMed
Google Scholar
Friend DJC (1965) Длина колоса и количество колосков пшеницы, выращенной при разных температурах и яркости света.Can J Bot 43: 345–353
Статья
Google Scholar
Гамбин Б.Л., Боррас Л. (2010) Распределение ресурсов и соотношение между количеством семян и массой семян: сравнение по видам сельскохозяйственных культур. Ann Appl Biol 156: 91–102
Статья
Google Scholar
Gawroński P, Ariyadasa R, Himmelbach A, Poursarebani N, Kilian B, Stein N, Steuernagel B, Hensel G, Kumlehn J, Sehgal SK (2014) Искаженные циркадные часы вызывают раннее цветение и температурные колебания колоса развитие мутанта Eps-3Am пшеницы еинкорня.Генетика 196: 1253–1261
Статья
CAS
PubMed
PubMed Central
Google Scholar
Gonzalez FG, Miralles DJ, Slafer GA (2011) Выживаемость цветков пшеницы в зависимости от роста колоса перед цветением. J Exp Bot 62: 4889–4901
Статья
CAS
PubMed
Google Scholar
Гринвуд JR, Finnegan EJ, Watanabe N, Trevaskis B, Swain SM (2017) Новые аллели гена одомашнивания пшеницы Q обнаруживают множественные роли в росте и репродуктивном развитии.Разработка 144: 1959–1965
Статья
CAS
PubMed
Google Scholar
Griffiths S, Wingen L, Pietragalla J, Garcia G, Hasan A, Miralles D, Calderini DF, Ankleshwaria JB, Waite ML, Simmonds J, Snape J, Reynolds M (2015) Генетическое вскрытие размера зерен и количества зерен компромиссы в зародышевой плазме пшеницы CIMMYT. PLoS One 10: e0118847
Артикул
CAS
PubMed
PubMed Central
Google Scholar
Guo Z, Schnurbusch T (2015) Изменение плодородия цветков у гексаплоидной пшеницы, выявленное при удалении побегов.J Exp Bot 66: 5945–5958
Статья
CAS
PubMed
PubMed Central
Google Scholar
Guo Z, Slafer GA, Schnurbusch T (2016) Генотипические вариации в признаках плодовитости колоса и размере завязи как детерминантах выживаемости цветков и зерна у пшеницы. J Exp Bot 67: 4221–4230
Статья
CAS
PubMed
PubMed Central
Google Scholar
Guo Z, Chen D, Alqudah AM, Röder MS, Ganal MW, Schnurbusch T (2017) Полногеномный ассоциативный анализ 54 признаков выявил множественные локусы для определения плодовитости цветков пшеницы.Новый Фитол 214: 257–270
Артикул
CAS
PubMed
Google Scholar
Guo Z, Chen D, Röder M, Ganal M, Schnurbusch T (2018) Генетическое расчленение длительностей подфаз перед цветением во время развития репродуктивного колоса пшеницы. Завод J 95: 909–918
Артикул
CAS
Google Scholar
Hucl P, Fowler BJ (1992) Сравнение разветвленной колосовой пшеницы с сортами Neepawa и Hy320 по урожайности зерна и компонентам урожайности.Can J Plant Sci 72: 671–677
Статья
Google Scholar
Хуш Г.С. (2001) Зеленая революция: путь вперед. Nat Rev Genet 2: 815–822
Статья
CAS
PubMed
Google Scholar
Лоулор Д.В., Пол М.Дж. (2014) Взаимодействие источника / поглотителя лежит в основе урожайности: пример трегалозо-6-фосфата / SnRK1 в улучшении урожая пшеницы. Фасадный завод Sci 5: 418
Артикул
PubMed
PubMed Central
Google Scholar
Льюис С., Фаричелли М.Э., Аппендино М.Л., Валарик М., Дубковски Дж. (2008) Хромосомная область, включая локус ранней ранней стадии как таковой Eps-Am1, влияет на продолжительность ранних фаз развития и количество колосков у диплоидной пшеницы.J Exp Bot 59: 3595–3607
Статья
CAS
PubMed
PubMed Central
Google Scholar
Li H (2011) Статистическая основа для вызова SNP, обнаружения мутаций, сопоставления ассоциаций и оценки генетических параметров популяции на основе данных секвенирования. Биоинформатика 27: 2987–2993
Статья
CAS
PubMed
PubMed Central
Google Scholar
Li H (2013) Выравнивание считываний последовательностей, последовательностей клонов и контигов сборки с помощью BWA-MEM.arXiv препринт. arXiv: 1303.3997
Li H, Handsaker B, Wysoker A, Fennell T, Ruan J, Homer N, Marth G, Abecasis G, Durbin R, Proc GPD (2009) Формат выравнивания / карты последовательностей и SAMtools. Биоинформатика 25: 2078–2079
Статья
CAS
PubMed
PubMed Central
Google Scholar
Лю Б., Лю Л., Тянь Л., Цао В., Чжу Ю., Ассенг С. (2014) Тепловой стресс после уборки урожая и влияние урожайности озимой пшеницы в Китае.Glob Change Biol 20: 372–381
Статья
Google Scholar
Martin M (2011) Cutadapt удаляет последовательности адаптеров из операций чтения с высокой пропускной способностью. EMBnet J 17: 10–12
Статья
Google Scholar
Martinez-Barajas E, Delatte T, Schluepmann H, de Jong GJ, Somsen GW, Nunes C, Primavesi LF, Coello P, Mitchell RA, Paul MJ (2011) Развитие зерна пшеницы характеризуется значительным содержанием трегалозы-6-фосфата. накопление наполнения перед зерном: распределение в тканях и связь с активностью протеинкиназы-1, связанной с SNF1.Plant Physiol 156: 373–381
Статья
CAS
PubMed
PubMed Central
Google Scholar
Mascher M, Wu SY, St Amand P, Stein N, Poland J (2013) Применение генотипирования путем секвенирования на платформах полупроводникового секвенирования: сравнение генетического и контрольного порядка маркеров в ячмене. PLoS One 8: e76925
Артикул
CAS
PubMed
PubMed Central
Google Scholar
McMaster GS (1997) Фенология, развитие и рост пшеницы ( Tritimm aestivum L.) стрелять апексом: обзор. В: Sparks DL (ed) Достижения в агрономии. Academic Press Inc, США, стр. 63–118. http://www.apnet.com
Google Scholar
McSteen P (2009) Гормональная регуляция ветвления у трав. Plant Physiol 149: 46–55
Статья
CAS
PubMed
PubMed Central
Google Scholar
Miralles DJ, Slafer GA (2007) Ограничения поглощения урожая пшеницы: как их можно уменьшить? J Agric Sci 145: 139–149
Статья
Google Scholar
Myers SS, Zanobetti A, Kloog I, Huybers P, Leakey AD, Bloom AJ, Carlisle E, Dietterich LH, Fitzgerald G, Hasegawa T, Holbrook NM, Nelson RL, Ottman MJ, Raboy V, Sakai H, Sartor KA, Schwartz J, Seneweera S, Tausz M, Usui Y (2014) Увеличение CO 2 угрожает питанию человека.Nature 510: 139–142
Статья
CAS
PubMed
PubMed Central
Google Scholar
Pennell A, Halloran GM (1984a) Влияние яровизации и фотопериода на экспрессию избыточных колосков у пшеницы. Ann Bot 53: 821–831
Статья
Google Scholar
Пеннелл А.Л., Халлоран Г.М. (1984b) Влияние времени посева, фотопериода и температуры на избыточную экспрессию колосков у пшеницы ( Triticum ).Can J Bot Revue Canadienne De Botanique 62: 1687–1692
Google Scholar
Персиваль Дж. (1921) Пшеничное растение: монография. Duckworth and Co., Лондон, стр. 241–261
Google Scholar
Pinthus MJ, Millet E (1978) Взаимодействие между количеством колосков, количеством зерен и массой зерна в колосьях пшеницы ( Triticum-Aestivum L). Ann Bot 42: 839–848
Статья
Google Scholar
Польша JA, Brown PJ, Sorrells ME, Jannink JL (2012) Разработка генетических карт высокой плотности для ячменя и пшеницы с использованием нового метода двухферментного генотипирования путем секвенирования.PLoS One 7: e32253
Артикул
CAS
PubMed
PubMed Central
Google Scholar
Poursarebani N, Seidensticker T, Koppolu R, Trautewig C, Gawronski P, Bini F, Govind G, Rutten T, Sakuma S, Tagiri A, Wolde GM, Youssef HM, Battal A, Ciannamea S, Fusca T, Nussbaumer T, Pozzi C, Borner A, Lundqvist U, Komatsuda T, Salvi S, Tuberosa R, Uauy C, Sreenivasulu N, Rossini L., Schnurbusch T. (2015) Генетическая основа составной формы колоса у ячменя и «чудо-пшеницы».Генетика 201: 155–165
Статья
CAS
PubMed
PubMed Central
Google Scholar
Rawson HM (1970) Число колосков, его контроль и связь с урожайностью на колос у пшеницы. Austr J Biol Sci 23: 1–16
Статья
Google Scholar
Rawson H (1971) Верхний предел количества колосков на колос у пшеницы, контролируемый фотопериодом. Crop Pasture Sci 22: 537–546
Статья
Google Scholar
Rawson HM, Richards RA (1993) Влияние высокой температуры и фотопериода на развитие цветков в изолиниях пшеницы, различающихся генами яровизации и фотопериода.Полевые культуры Res 32: 181–192
Статья
Google Scholar
Rawson H, Ruwali K (1972) Колосовое ветвление как средство повышения однородности зерна пшеницы. Aust J Agric Res 23: 551–559
Статья
Google Scholar
Рейнольдс М., Кальдерини Д., Кондон А., Варгас М. (2007) Ассоциация признаков источника / поглотителя с урожайностью, биомассой и эффективностью использования излучения среди случайных сестринских линий от трех кроссов пшеницы в среде с высокой урожайностью.J Agric Sci 145: 3–16
Статья
Google Scholar
Reynolds M, Foulkes MJ, Slafer GA, Berry P, Parry MAJ, Snape JW, Angus WJ (2009) Повышение потенциала урожайности пшеницы. J Exp Bot 60: 1899–1918
Статья
CAS
PubMed
Google Scholar
Ричардс Р. (1996) Повышение урожайности пшеницы: управление источниками и поглотителями. В: Рейнольдс М.П., Раджарам С., Макнаб А. (ред.) Повышение урожайности пшеницы: преодоление барьеров.Международный центр улучшения кукурузы и пшеницы, Сонора, стр 134–149
Google Scholar
Röder MS, Korzun V, Wendehake K, Plaschke J, Tixier MH, Leroy P, Ganal MW (1998) Микросателлитная карта пшеницы. Генетика 149: 2007–2023
PubMed
PubMed Central
Google Scholar
Ройо С., Альваро Ф., Мартос В., Рамдани А., Исидро Дж., Виллегас Д., дель Мораль LFG (2007) Генетические изменения в компонентах урожая твердой пшеницы и связанных с ними чертах у итальянских и испанских сортов в ХХ веке.Euphytica 155: 259–270
Статья
Google Scholar
Санчес-Гарсия М., Ройо С., Апарисио Н., Мартин-Санчес Дж. А., Альваро Ф. (2013) Генетическое улучшение урожайности мягкой пшеницы и связанных с ней признаков в Испании в 20 веке. J Agric Sci 151: 105–118
Статья
CAS
PubMed
Google Scholar
Сато-Нагасава Н., Нагасава Н., Малкомбер С., Сакаи Х., Джексон Д. (2006) Фермент метаболизма трегалозы контролирует структуру соцветий кукурузы.Nature 441: 227–230
Статья
CAS
PubMed
Google Scholar
Серраго Р.А., Альзуэта И., Савин Р., Слафер Г.А. (2013) Понимание реакции урожайности зерна на соотношение источник-поглотитель во время заполнения зерна пшеницей и ячменем в контрастных средах. Полевые культуры Res 150: 42–51
Статья
Google Scholar
Шарман Б (1944) Ветвистые головки у пшеницы и гибридов пшеницы.Nature 153: 497–498
Статья
Google Scholar
Шоу Л.М., Тернер А.С., Херри Л., Гриффитс С., Лори Д.А. (2013) Мутантные аллели фотопериода-1 у пшеницы ( Triticum aestivum L.), которые придают фенотип позднего цветения в течение долгих дней. PLoS One 8: e79459
Артикул
CAS
PubMed
PubMed Central
Google Scholar
Slafer GA, Miralles DJ (1993) Эффективность плодоношения трех сортов мягкой пшеницы ( Tritkum aestivum ), выпущенных в разные эпохи.Количество зерен в колосе и масса зерен. J Agron Crop Sci 170: 251–260
Статья
Google Scholar
Слафер Г.А., Савин Р. (1994) Взаимосвязь между источником и стоком и масса зерна в различных местах в пределах колоса пшеницы. Полевые культуры Res 37: 39–49
Статья
Google Scholar
Slafer GA, Elia M, Savin R, García GA, Terrile II, Ferrante A, Miralles DJ, González FG (2015) Эффективность плодоношения: альтернативный признак для дальнейшего повышения урожайности пшеницы.Food Energy Secur 4: 92–109
Статья
Google Scholar
Соссо Д., Луо Д., Ли К.Б., Сассе Дж., Ян Дж., Гендрот Дж., Сузуки М., Кох К.Э., Маккарти Д.Р., Чоури П.С., Роговски П.М., Росс-Ибарра Дж., Ян Б., Фроммер В.Б. (2015) Засыпка семян домашней кукурузы и риса зависит от транспорта гексозы, опосредованного SWEET. Nat Genet 47: 1489
Статья
CAS
PubMed
Google Scholar
Международный консорциум по секвенированию генома пшеницы (IWGSC) (2014) Проект последовательности генома гексаплоидной мягкой пшеницы ( Triticum aestivum ) на основе хромосом.Science 345: 1251788
Статья
CAS
Google Scholar
Thomas SG (2017) Novel Rht-1 карликовые гены: инструменты для селекции пшеницы и анализа функции белков DELLA. J Exp Bot 68: 354–358
Статья
CAS
PubMed
PubMed Central
Google Scholar
USDA (1916) Аляска и стоунер, или «чудо», пшеница: две разновидности сильно искажены.Бюллетень № 357, Министерство сельского хозяйства США, Вашингтон, округ Колумбия
White AC, Rogers A, Rees M, Osborne CP (2016) Как мы можем заставить растения расти быстрее? Взгляд источника-поглотителя на темпы роста. J Exp Bot 67: 31–45
Статья
CAS
PubMed
Google Scholar
Уорланд А., Бёрнер А., Корзун В., Ли В., Петрович С., Сэйерс Э. (1998) Влияние генов фотопериода на адаптивность европейских озимых пшениц.Euphytica 100: 385–394
Статья
CAS
Google Scholar
Ян Л., Лукоянов А., Транквилли Г., Хельгуера М., Фахима Т., Дубковский Дж. (2003) Позиционное клонирование гена яровизации пшеницы VRN1. Proc Natl Acad Sci 100: 6263–6268
Статья
CAS
PubMed
Google Scholar
Yan L, Loukoianov A, Blechl A, Tranquilli G, Ramakrishna W, SanMiguel P, Bennetzen JL, Echenique V, Dubcovsky J (2004) Ген пшеницы VRN2 — репрессор цветения, подавляемый яровизацией.Science 303: 1640–1644
Статья
CAS
PubMed
PubMed Central
Google Scholar
Yan L, Fu D, Li C, Blechl A, Tranquilli G, Bonafede M, Sanchez A, Valarik M, Yasuda S, Dubcovsky J (2006) Ген яровизации пшеницы и ячменя VRN3 является ортологом FT. Proc Natl Acad Sci 103: 19581–19586
Статья
CAS
PubMed
Google Scholar
Youssef HM, Eggert K, Koppolu R, Alqudah AM, Poursarebani N, Fazeli A, Sakuma S, Tagiri A, Rutten T., Govind G, Lundqvist U, Graner A, Komatsuda T, Sreenivasulu N, Schnurbusch T, 2017 ) VRS2 регулирует опосредованное гормонами формирование рисунка соцветий ячменя.Nat Genet 49: 157–161
Статья
CAS
Google Scholar
Youssefian S, Kirby EJM, Gale MD (1992a) Плейотропные эффекты GA-нечувствительных генов карликовости Rht в пшенице. 1. Воздействие на развитие колоса, стебля и листьев. Полевые культуры Res 28: 179–190
Статья
Google Scholar
Youssefian S, Kirby EJM, Gale MD (1992b) Плейотропные эффекты GA-нечувствительных генов карликовости Rht в пшенице.2. Воздействие на рост листьев, стеблей, початков и цветков. Полевые культуры Res 28: 191–210
Статья
Google Scholar
Yu SM, Lo SF, Ho TH (2015) Связь между источником и стоком: регулируется перекрестной передачей сигналов гормонов, питательных веществ и стресса. Trends Plant Sci 20: 844–857
Статья
CAS
PubMed
Google Scholar
Чжан Х., Тернер NC, Пул М.Л. (2010) Баланс «источник – поглотитель» и управление отношениями «поглотитель – источник» пшеницы указывают на то, что потенциал урожайности пшеницы ограничен поглотителем в зонах с большим количеством осадков.Crop Pasture Sci 61: 852–861
Статья
Google Scholar
Zikhali M, Wingen LU, Griffiths S (2016) Определение границ гена раннего цветения per se D1 ( Eps-D1 ) до субтеломерной хромосомной делеции в мягкой пшенице ( Triticum aestivum ). J Exp Bot 67: 287–299
Статья
CAS
PubMed
Google Scholar
семейств Poaceae — Растения Огайо
POACEAE
(семейство трав)
Как и другие основные грамиоиды — осоки семейства Cyperaceae — травы представляют собой однодольные с линейными листьями и крошечными цветками, опыляемыми ветром.В отличие от осоки, принципиальное значение которой имеет экологическое значение, поскольку она приносит пользу животным водно-болотных угодий, травы являются важными как важными составляющими очень многих природных экосистем, так и экономически, поскольку семейство включает в себя основные пищевые растения большинства цивилизаций в виде растений. рис, пшеница, кукуруза (кукуруза), рожь, овес, ячмень, просо и сорго. Этот травяной плод представляет собой «зерно», также называемое «зерновкой», то есть односемянный нерасцветающий сухой плод, очень похожий на семянку, за исключением того, что стенка завязи зерна плотно сращена с семенем и не может быть отделена от нее, кроме измельчение (например, когда белый рис сделан из коричневого риса).
Чтобы отличить обычную траву от осоки, запомните броский слоган, который помогает отличить травы от осоки: « У травы нет краев ». Типичный стебель травы в поперечном сечении круглый. Кроме того, часть основания листа, которая огибает стебель осоки, называемая влагалищем листа, у большинства трав открыта, то есть имеет свободные края, один из которых перекрывает другой. В отличие от этого, у осок оболочка закрыта, а это означает, что она непрерывна вокруг стебля на стороне, противоположной листовой пластинке.
У травы нет краев!
У злаков есть крошечные, опыляемые ветром цветки без лепестков и чашелистиков. Они окружены специальными чешуйчатыми прицветниками и собраны в особый тип соцветий, называемый «колоском» (крошечный колос). Колоски травы сплюснутые, с цветами, прикрепленными двумя противоположными рядами, и с несколько замысловатым расположением, которое включает дополнительные чешуевидные прицветники, встречающиеся только у трав.В отличие от этого, колоски осоки часто (но не всегда) состоят из цветов, расположенных плотной спиралью, внутри которой каждый цветок заключен в одну чешуйку.
Цветки злаков крошечные, опыляются ветром, расположены в виде колосков.
Коллекция цветков и специализированных прицветников, составляющих колоск травы, обычно состоит из пары «чешуек» у самого основания колосков, над которыми расположены один или несколько «цветочков», каждый из которых состоит из рудиментарного цветка, окруженного пара чешуевидных прицветников.Это большая, самая внешняя «лемма», которая охватывает меньшую, самую внутреннюю «палею».
Чешуйки и чешуйки, о боже!
Цветки, такие как они есть, в основном состоят из завязи с короткой вершиной и пары перистых рыльц, которые отлично улавливают переносимую ветром пыльцу, плюс 2 или 3 тычинки, состоящие из пропорционально больших пыльников на конце длинных тонкие свисающие нити.
Колосок травы состоит из пары чешуек, над которыми расположены один или несколько цветков.
Во время их короткого периода цветения очевидны перистые рыльца, превосходно приспособленные для приема летающих по воздуху пыльцевых зерен. В дополнение к этому, тычинки с их большими пыльниками и очень тонкими нитями отлично переносят пыльцу на ветер. Вот, например, цветет просо.
Цветки просо проса имеют оранжевые пыльники и пурпурные рыльца.
В то время как колоски являются типом соцветий трав, сами колоски расположены во вторичных соцветиях некоторого типа.Вот несколько примеров.
Просо дает диффузную метелку.
Просо дает диффузную метелку.
Quackgrass и итальянский райграс дают колоски на удлиненном шипе, но с интересной разницей. Колоски кряквы расположены стороной к стеблю, а колоски райграса — сбоку к стеблю.
Слева: колоски кряквы сбоку к стеблю .
Справа: колосья ржаной травы на боку.
НЕСКОЛЬКО РАСПРОСТРАНЕННЫХ ТРАВ
В мае поищите несколько интродуцированных, даже сорных трав, которые выделяются вдоль обочин. Род Bromus (бром) включает виды с многоцветковыми колосками, расположенными в метелках. У некоторых видов чешуйки заметно остистые. Виды схожи и разделяют сорняковую экологию, поэтому на данный момент их можно было бы назвать «остистым костром».”
Трава брамовника остистая сорная, с крупными колосками на рыхлых метелках.
Овсяница высокорослая, Festuca pratensis (ранее известная как Festuca elatior ) — это прочное многолетнее растение, которое дает несколько цветков, нераспустившихся колосков на вертикальной метелке.
Овсяница высокорослая дает метелку из нераспустившихся колосков с узкими цветками.
Райграс английский (не итальянский), Lolium perenne дает несколько цветковых колосков в виде колоса, которые прикрепляются к рахису по бокам
Колоски английского райграса расположены в виде колоса и расположены сбоку от стебля.
Фруктовая трава, Dactlyis glomerata , дает малоцветковые колоски в метелке, при этом они скучены к концам ветвей метелки и выглядят как маленькие кулаки.
Пятна садовой травы слипаются к концам ветвей метелки.
Контроль судьбы меристемы колосков кукурузы с помощью APETALA2-подобного гена undeterminate spikelet1
- Джордж Чак,
- Роберт Б.Мили и
- Сара Хейк
- Департамент биологии растений и микробов, Калифорнийский университет, Беркли, Калифорния , США;
Pioneer Hi-Bred International, Джонстон, Айова 50131, США;
Центр экспрессии генов растений, Служба сельскохозяйственных исследований Министерства сельского хозяйства США (USDA-ARS), Олбани, Калифорния ,
США
Абстрактные
Упорядоченное производство меристем с определенной судьбой имеет решающее значение для правильной разработки архитектуры растений.Кукуруза
меристема соцветия разветвляется несколько раз, образуя боковые меристемы с определенными судьбами. Образовалась первая меристема,
меристема пары колосков дает две меристемы колосков, каждая из которых дает две цветочные меристемы. Мы определили
ген, названный undeterminate spikelet1 ( ids1 ), который определяет детерминированную судьбу меристемы колосков и тем самым ограничивает количество продуцируемых цветочных меристем.В отсутствие
Из-за функции гена ids1 меристема колосков становится неопределенной и дает дополнительные соцветия. Члены семейства злаковых различаются
по количеству цветков в их колосках, предполагая, что ids1 могут играть роль в архитектуре соцветий у других видов трав. ids1 является членом семейства генов транскрипционных факторов APETALA2 ( AP2 ), которые участвуют в широком диапазоне ролей в развитии растений.Выражение
ids1 был обнаружен во многих типах зачатков боковых органов, а также в меристемах колосков. Наш анализ мутантного фенотипа ids1, и паттерна экспрессии показывает, что ids1 определяет детерминированные судьбы путем подавления неопределенного роста в меристеме колосков.
Развитие тела растения зависит от активности апикальных меристем, групп неопределенных клеток, обнаруженных на
советы по выращиванию.Способность апикальных меристем побегов оставаться неопределенными позволяет им постоянно инициировать органы,
ткани и вторичные меристемы, необходимые для нормального развития. Это свойство требует, чтобы меристема выделяла популяцию.
клеток, чтобы обновиться и восполнить потери клеток в каждом боковом органе или вторичной меристеме. Провал этого процесса
произойти приводит к прекращению роста кончика. Некоторые меристемы, например цветочные меристемы, потребляются при производстве
боковых органов и могут быть описаны как имеющие определенную судьбу.Соцветие кукурузы особенно полезно для
изучение детерминированности меристемы, поскольку она претерпевает несколько четко определенных событий ветвления, чтобы произвести меристемы со все более
ограниченные судьбы.
Соцветия травы организованы в группы, называемые колосками, каждый из которых состоит из пары стерильных прицветников, называемых чешуйками, покрывающими
фиксированное количество цветков (Clifford 1987). Регулирование количества цветков на колоске является основным фактором, определяющим архитектуру колосков среди членов травы.
семья.Колосья кукурузы детерминированное, дает только два цветочка в определенных положениях на оси, называемой рахиллой.
(Weatherwax 1923). Родственники кукурузы характеризуются колосками с неопределенным количеством цветков, например пшеница, или содержат только
по одному цветочку на колоск, как у ячменя. Один из классических критериев, используемых для различения различных видов трав, заключается в том, являются ли они
колоски содержат определенное или неопределенное количество цветков (Clifford and Watson 1977).Сам колоск — лишь один из компонентов сложного разветвленного соцветия кукурузы. В отличие от двудольных
у таких растений, как Arabidopsis , у которых цветки образуются непосредственно апикальной и боковой меристемами (Hempel and Feldman 1994), у кукурузы есть по крайней мере две отчетливые стадии ветвления соцветий до того, как меристема колосков завершится в производстве.
из двух цветочков. Эти дополнительные ступени ветвления обеспечивают большее морфологическое разнообразие трав.
У кукурузы был описан ряд мутантов, которые приводят к появлению дополнительного количества цветков внутри колоска (Veit et al. 1993). У разветвленных бесшелковых мутантов дополнительные соцветия начинаются у мужских колосков кисточки (Kempton 1934), хотя более радикальная трансформация наблюдается в женских колосках уха, у которых цветочки превращаются в длинные
индетерминантные ветви (Veit et al. 1993). Исследования мутации Tasselseed6 показали, что меристема колосков задерживается в достижении детерминированности, что позволяет ей запускать соцветия.
в течение более длительного периода времени (Irish et al.1994). Анализ мутантов Tasselseed6 и привел к модели (Irish 1997), в которой меристема соцветия и ее ответвления проходят через упорядоченную, определенную серию детерминированных онтогенетических групп.
состояний, заканчиваясь превращением терминальной меристемы колосков в верхний цветочек. Подобные ветвящиеся мутанты имеют
также были описаны для других видов трав и включают мутацию многоцветкового ячменя (Bossinger et al., 1992) и доминантный мутант Naked овса (Ougham et al.1996).
Множество генетических и молекулярных исследований выявили несколько генов, важных для развития цветков. Один такой ген,
гомеотический ген APETALA2 ( AP2 ) из Arabidopsis, выполняет несколько функций в развитии цветов, семян и семяпочек (Kunst et al. 1989; Jofuku et al. 1994; Modrusan et al. 1994). Помимо своей роли в определении идентичности цветочных органов, AP2 влияет на регуляцию идентичности цветочных меристем.Например, двойные мутанты слабого аллеля ap2-1 с мутантами идентичности цветочной меристемы, такими как листовой или apetala1 , дают больше боковых ветвей соцветия вместо цветков (Bowman et al. 1993). Кроме того, слабые аллели ap2 в короткие дни вызывают образование третичных цветочных побегов в пазухах трансформированных чашелистиков (Schultz and Haughn 1993). Ген AP2 принадлежит к большому семейству генов, 12 из которых были идентифицированы у Arabidopsis (Okamuro et al.1997). Многочисленные гомологи были идентифицированы как у однодольных, так и у двудольных (Jofuku et al. 1994; Ohme-Takagi and Shinshi 1995; Moose and Sisco 1997). Мутации в AP2 -подобном гене, AINTEGUMENTA, являются дефектными в развитии яйцеклетки (Elliot et al. 1996; Klucher et al. 1996). Недавно было показано, что ген glossy15 кукурузы является геном, подобным AP2 , который репрессирует признаки взрослых листьев в молодых листьях (Moose and Sisco 1997).
Здесь мы описываем новый мутант ветвления кукурузы, undeterminate spikelet1 ( ids1 ), который мы получили путем анализа фенотипа, обусловленного потерей функции гена, подобного AP2 кукурузы. ids1 мутанты имеют неопределенный колоск, в котором образуется несколько цветков вместо двух, обнаруженных у кукурузы дикого типа.
ids1 Экспрессия наблюдалась во множестве боковых органов, а также в паре колосков и меристемах колосков. Наш анализ
указывает на то, что ген ids1 является критическим для регуляции детерминированности меристемы колосков кукурузы.
Результаты
Выделение гена
ids1
Ген AP2 из Arabidopsis был использован для скрининга двух библиотек кДНК кукурузы с низкой строгостью, одна получена из незрелых початков, а другая — из вегетативных.
меристемы.Один и тот же класс кДНК был выделен из каждой библиотеки. Самый длинный клон этого класса состоял из 1967 нуклеотидов и
содержала ORF из 433 аминокислот (фиг.) с несколькими доменами, демонстрирующими поразительное аминокислотное сходство с геном AP2 из Arabidopsis (Jofuku et al. 1994). Были обнаружены два тандемно повторяющихся мотива из 68 аминокислот, которые на 86% идентичны аминокислотам с доменом AP2 белка Arabidopsis AP2. Семейство гена AP2 можно разделить на две группы, обозначенные как EREBP-подобные или AP2-подобные, в зависимости от того, обладают ли они
один или два повтора AP2 соответственно (Okamuro et al.1997). Белок IDS1 относится к последнему классу, который включает белки AP2, AINTEGUMENTA, GLOSSY15 и RAP2.7 (рис.)
(Jofuku et al. 1994; Klucher et al. 1996; Moose and Sisco 1997; Okamuro et al. 1997). Белки табака ERE-BP, которые имеют только один из этих повторов, связывают ДНК (Ohme-Takagi and Shinshi 1995). Таким образом, по аналогии вполне вероятно, что IDS1 функционирует как фактор транскрипции. В подтверждение этого краткий отрезок основных
аминокислоты, которые могут функционировать как домен ядерной локализации, присутствуют в белке IDS между 100 и 110 аминокислотами.
(Рисунок.). Белок AP2 из Arabidopsis содержит богатый серином кислотный домен на аминоконце, который может функционировать как домен активации (Jofuku et al. 1994). Хотя подобный участок находится на аминоконце белка IDS1 в положениях 9–45, этот участок намного меньше
и имеет меньше кислотных и сериновых остатков по сравнению с AP2. За пределами домена AP2 наблюдается очень небольшое сходство последовательностей
между AP2 и IDS1. Учитывая это открытие, удивительно, что положения шести интронов в домене AP2 сохраняются.
между ids1 и AP2 (рис.). Аналогичный результат был получен для гена gl15 кукурузы (Moose and Sisco 1997).
Нуклеотид и выведенная аминокислотная последовательность IDS1. Аминокислотные числа показаны слева от ; нуклеотидов пронумерованы справа . Подчеркнутые аминокислотные последовательности представляют два домена AP2 (Jofuku et al.1994). Серины и кислые аминокислоты на амино-конце обозначены коротким подчеркиванием. Аминокислотные последовательности выделены жирным шрифтом
представляют собой основной регион с предполагаемой активностью ядерной локализации. Жирные нуклеотидные последовательности соответствуют ZAP2-1.
праймер, используемый для скрининга ПЦР. (▿) шесть положений интронов между IDS, AP2, и GL15 сохраняются; (▾) другой интрон.
Сравнение между доменами AP2 AP2 -подобных генов.Выведенные аминокислотные последовательности доменов AP2 IDS1 (номер доступа в GenBank AF048900), RAP2.7 (AF003100), GL15 (U41466) и ANT (U40256) сравниваются с AP2 (ATU12546). Область между двумя повторами AP2, то есть линкерная область, начинается с аминокислоты
78 и заканчивается на 102. Для облегчения совмещения были введены зазоры, представленные точками. Аминокислоты, общие для всех пяти
белки выделены жирным шрифтом и в согласованной последовательности внизу .
ids1 был картирован в длинном плече первой хромосомы в положении 192 с использованием рекомбинантных инбредных линий (Burr et al. 1988), зондированных неповторяющимся фрагментом ДНК с 3′-конца ids1. Никакие известные морфологические мутанты не были картированы в этой позиции.
Выделение рецессивных
ids1 мутаций
Для выяснения функции ids1, мы использовали обратную генетическую стратегию для генерации аллелей с потерей функции. ПЦР использовали на большой популяции растений, несущих
Мутатор транспозонов ( Mu ) для поиска вставок в ids1 (Bensen et al.1995; Мили и Бриггс 1995). Праймеры, фланкирующие домен AP2 ids1 в комбинации с праймерами Mu , дали два независимых события вставки через этот скрининг. Секвенирование продуктов ПЦР позволило локализовать вставки
точнее. Как показано на рисунке, вставка Mu в ids1-mum1 была локализована в пятом консервативном интроне домена AP2, тогда как вставка ids1-mum2 была локализована в первом консервативном интроне на расстоянии 6 п.н. акцепторный сайт сплайсинга.Поскольку Mu элементы генерируют дупликацию 9 пар оснований при вставке (Bennetzen et al. 1993), акцепторный сайт сплайсинга также обнаруживается на 5′-конце Mu.
Локализация элементов Mu в ids1 – mum1 и ids1 – mum2. ids1 Последовательности интронов и экзонов показаны нормальным и жирным шрифтом соответственно.Номера интронов и экзонов показаны выше.
последовательности ДНК. Подчеркнутые последовательности представляют характерную дупликацию 9 п.н., связанную со вставками Mu .
Линии, несущие аллели ids1 – mum1 и ids1 – mum2 , демонстрировали рецессивный цветочный фенотип, который был более серьезным в линиях ids1 – mum2 .Чтобы убедиться, что вставки в ids1 ответственны за цветочный фенотип, мы проанализировали ДНК из 104 растений F2, используя ген ids1 в качестве зонда на гель-блотах ДНК. Косегрегация новых RFLP с цветочным фенотипом наблюдалась для 29 хромосом.
содержащий аллель ids1-mum1 и 91 хромосому, содержащую ids1-mum2 (данные не показаны). Не было обнаружено рекомбинантов между цветочным фенотипом и ids1-mum -специфическими полиморфизмами ни для одного из аллелей.
Мы проанализировали фенотипы ids1 – mum растений после обратного скрещивания с A632 и самоопыления. Наиболее устойчивым фенотипом у мутантов ids1 – mum был дефект в колоске. Зрелые колоски кисточки дикого типа содержат два цветочка, каждый из которых состоит из трех тычинок.
и две лодикулы, заключенные в прицветники палеи и леммы (рис. B). Палеа отличается от леммы в силу того, что
его положение и тот факт, что он двухкилочный с двойной средней жилкой (Clifford and Watson 1977).Колоски кисточек ids1 – mum2 больше, чем у кистей дикого типа (Рис. A), из-за того, что присутствуют лишние соцветия (Рис. C). Номер
Количество цветков колеблется от минимум 3 из ids1 – mum1 растений до 10 из ids1 – mum2 растений. Помимо этих соцветий, между ними обычно можно увидеть тонкую рахиллу, содержащую несколько более мелких соцветий.
два самых верхних цветочка (рис. F). Эти более мелкие соцветия, как правило, постепенно уменьшаются, так что оставшиеся
кончик рахиллы едва заметен.Более крупные дополнительные соцветия показывают нормальный образец половой дифференциации и
имеют правильное количество тычинок и лодикул, окруженных палеей и леммой (рис. C).
Фенотипы у ids1 – мама мужских и женских колосков. ( A ) Колосок с кисточкой нормальный A632 ( левый, ) и ids1 – mum2 колосок с кисточкой ( правый ).( B ) Рассеченный нормальный колоск кисточки A632, показывающий боковые органы верхних и нижних соцветий. ( C ) Расщепленный колоск с кисточкой ids1 – mum2 с четырьмя цветками вместо двух. У каждого цветочка по три тычинки, окруженные палеой и леммой, как в A632.
( D ) Ушной колосок нормальный A632 ( левый ) и ids – mum1 ушной колоск ( правый ). Стрелки указывают на шелк. ( E ) Срединные продольные срезы неоплодотворенного 25-дневного ребенка ids1 – mum2 ушной колоск ( левый ) и ушной колоск A632 ( правый ).Черные стрелки указывают на цветочки. Колоски А632 имеют одиночный цветочек с увеличенным нуцеллусом; ids1 – mum2 ушные колоски имеют по крайней мере три цветочка со значительно сниженным развитием боковых органов. ( F ) ids1 – mum2 колоск на ухе с удлиненной рахиллой, содержащей два цветочка. Одиночная стрелка указывает на кончик рахиллы.
(st) тычинки; (il) внутренняя лемма; (па) палеа; (ol) внешняя лемма; (ig) внутренняя чешуйка; (og) наружная чешуйка; (уф) верхний цветочек; (lf) ниже
цветочек; (ra) рахилла; (фл) цветочек.
Колоски самок ids1 – mum также имеют больший размер из-за наличия лишних цветков (рис. D, E). Колоски нормальные женские
содержат только один зрелый цветочек из-за аборта нижнего цветочка (Делонг и др., 1993). Развитие вторичных половых признаков у женских цветков включает в себя прерывание тычинок и развитие
гинецея, на примере образования длинных шелков, состоящих из двух сросшихся плодолистиков (Randolph 1936). ids1 – mum1 колоски самок часто содержат более одного шелка, расположенного в эктопических положениях (Рис. D). Эти шелка, однако, обычно
не развиваются нормально; они часто не сливаются и не удлиняются до нужной длины. Если проявляющееся ухо вручную
После вскрытия и опыления развиваются несколько зерен, что позволяет предположить, что все другие аспекты оплодотворения и развития плодов
обычный. Достигнув зрелости, эти ядра демонстрируют ids1 – mum гомозиготных фенотипов, и поэтому не представляют собой изъятия из зародыша элементов Mu .Аллель ids1 – mum2 кажется более серьезным, поскольку большинство лишних цветков не могут развить полностью сформированные боковые органы (Рис. E, слева).
Приблизительно 5% из ids1 – mum2 женских колосков имеют удлиненную рахиллу, выходящую из колоска (Рис. F). В таких случаях можно заметить, что
рахилла не оканчивается верхним цветком, что еще раз подтверждает ее неопределенный характер. Нет очевидного и последовательного
фенотипы наблюдались в листьях и корнях мутантов ids1, , несмотря на то, что ids1 экспрессируется в этих органах.
Анализ с помощью растровой электронной микроскопии
Сканирующая электронная микроскопия использовалась для определения точки, в которой развитие меристемы колосков ids1 – mum отличается от развития меристемы колосков дикого типа. Меристема соцветия обычно инициирует пару колосков.
меристемы акропетально, которые, в свою очередь, разветвляются, образуя две колосковые меристемы (рис.А) (Ченг и др., 1983). Меристемы колосков также разветвляются, образуя нижние и верхние соцветия. Эти цветочки образуют необычный узор,
то есть, чередуя 180 градусов друг от друга (Рис. B). Хотя нижняя цветочная меристема инициирует сначала, боковой орган
В верхнем цветке развитие происходит значительно раньше, чем в нижнем (рис. C, D). Палеа — первый боковой орган
чтобы дифференцироваться, и вскоре после этого развиваются три тычинки (Cheng et al.1983). Половая дифференциация происходит на более поздних стадиях, когда гинецей прерывается у мужских цветков, а тычинки — у самок.
цветочки (Dellaporta, Calderon-Urrea, 1993).
Сканирующая электронная микроскопия нормальных и ids1 – mum2 ушей. ( A ) A632 зачаток уха с инициирующими зачатками пары колосков и зачатками колосков.Пруток, 300 мкм. ( B ) A632 неразветвленная меристема колосков ( верхняя ) и несколько более старая ветвистая меристема колосков ( низ ). Более старая меристема колосков подвергается боковому ветвлению, инициируя нижний цветочек. Бар, 102 мкм. ( C ) Развитие боковых органов у ушных колосков A632. В первую очередь развиваются боковые органы верхнего цветочка. Пруток, 80 мкм. ( D ) Созревающие боковые органы верхнего цветочка ушных колосков A632 с инициирующим гинецким гребнем.Бар, 104 мкм. ( E ) Ветвление ids1 – mum2 меристем колосков. Самая верхняя меристема колосков еще не разветвлена и выглядит удлиненной. Одновременное ветвление цветков
зарождение леммы происходит на противоположных сторонах средней и нижней меристем колосков. Штанга, 156 мкм. ( F ) Дальнейшее развитие ids1 – mum2 колосковых меристем с удаленными чешуйками, что свидетельствует о развитии цветочных меристем в пазухах нижних конечностей. Колоск
меристема сохраняется после каждого события ветвления.Пруток, 58 мкм. ( G ) Более старые ids1 – mum2 меристема колосков с удаленной чешуей. Остаточная меристема колосков удлиняется, образуя новый цветочек. Пруток, 60 мкм. ( H ) ids1 – mum2 колоск, показывающий созревающие боковые органы. Нет различия между верхним и нижним цветочками с точки зрения бокового органа.
разработка. Самый молодой цветочек, отходящий от меристемы колосков, не виден леммой. Бар, 171 мкм. (spm) Колоск
парная меристема; (см) колосковая меристема; (lfm) нижняя цветочная меристема; (gy) гинецей; (fm) цветочная меристема; (ле) лемма.
ids1 – mum мутанты обнаруживают нормальное развитие до стадии, на которой меристема колосков начинает ветвиться. На этом этапе меристемы колосков ids1 – mum кажутся слегка удлиненными (Рис. E, вверху). В несколько более старых меристемах колосков (рис. E, в середине) два отчетливых
события ветвления можно увидеть по наличию двух лемм (стрелок).Затем в пазухах этих пазух образуются цветочные меристемы.
леммы (рис. E, внизу и F). Эти события ветвления происходят на противоположных сторонах меристемы колосков по двоякой схеме.
Меристема колосков сохраняется после каждого события ветвления и остается между двумя последними сформированными цветочками (Рис. G, H). Этот
остаточная меристема колосков продолжает дистично инициировать дополнительные леммы и соцветия (рис. F, G). Скорость бокового органа
Развитие между первыми двумя ids1 – mum соцветий аналогично (рис.H), в отличие от дикого типа, у которого первыми развиваются органы верхнего цветочка.
Мы использовали гомеобоксный ген узлов, ( kn1 ) кукурузы, который экспрессируется в меристемах, а не в детерминированных боковых органах, для отслеживания событий ветвления.
в ids1 – мама колосков. kn1 экспрессируется в меристемах колосков и цветков кукурузы (рис. A, B) (Jackson et al. 1994).Гибридизация с антисмысловым зондом kn1 на меристемах колосков ids1-mum2 показала нормальную экспрессию в меристемах колосков (фиг. C). Однако, как только меристема колосков ids1-mum2 разветвляется с образованием цветков, меристема колосков становится больше и, следовательно, домен экспрессии kn1 расширяется (Рис. D). Сильная экспрессия kn1 наблюдается внутри дополнительных цветков, начинающихся от меристемы колосков на более поздних стадиях (рис.E, стрелки)
подтверждая меристематическое происхождение дополнительных цветков. Все другие аспекты экспрессии kn1 внутри цветков, включая отсутствие в боковых органах и слое L1, кажутся нормальными.
Локализация in situ kn1 в меристемах колосков A632 и ids1 – mum2 .( A ) Экспрессия kn1 внутри неразветвленной меристемы колоска A632. Пруток, 50 мкм. ( B ) Экспрессия kn1 внутри ветвящейся меристемы колоска A632. Выражение можно увидеть в зарождающемся нижнем цветочке. Пруток, 50 мкм.
( C ) Экспрессия kn1 в неразветвленной меристеме колосков ids1 – mum2 . Пруток, 50 мкм. ( D ) Экспрессия kn1 в пределах раннего ветвления ids1 – mum2 меристема колосков.По обе стороны от меристемы колосков видны зарождающиеся соцветия. Пруток, 60 мкм. ( E ) Экспрессия kn1 в более старом колоске ids1 – mum2 . Внутри развивающихся соцветий сохраняется сильное выражение. Пруток, 80 мкм. Стрелки указывают на зарождающиеся соцветия.
B, D, и E.
Анализ
ids1 выражения
Первоначальная характеристика экспрессии ids1 с использованием гель-блотов РНК показала, что ген экспрессируется как в вегетативных, так и в цветочных тканях.РНК-гель
блоттинг с 3′-частью кДНК ids1 показывает, что транскрипт ids1 присутствует во всех протестированных тканях (рис. A). Наименьшее количество экспрессии наблюдалось в ткани эмбриона, а наибольшее — в тканях эмбриона.
в ткани соцветия.
Анализ экспрессии ids1 в различных органах кукурузы B73.( A ) РНК-гель-блот, содержащий 1 мкг поли (A) + РНК, выделенной из эмбрионов через 17 дней после опыления ( E ), меристемы побегов, включая нерасширенный стебель и самые молодые зачатки листьев ( M ), колос примордии соцветия длиной до 2 см ( I ), первичные корни семян, проросших во влажном воздухе ( R ), и лопаточная часть полностью распустившихся молодых листьев ( L ) были промоканы и гибридизированы с 3 ‘частью. из кДНК ids1 , которая не включает домен AP2.( B ) Контрольная гибридизация. Блот в A зондировали кДНК убиквитина кукурузы, чтобы показать относительную нагрузку.
Для определения уровней транскрипта ids1 в мутантах ids1 – mum , поли (A) + РНК выделяли из мутантных ушей и сравнивали с РНК из ушей дикого типа. Как показано на рисунке А, транскрипт размера дикого типа отсутствует.
был обнаружен в аллеле ids1 – mum .Вместо этого наблюдались слабые транскрипты с более высокой молекулярной массой, которые могут быть результатом химерного или альтернативного
склеенные транскрипты. Этот результат демонстрирует, что вставки элементов Mu в ids1 не сплайсируются, и что как ids1-mum1 , так и ids1-mum2 могут представлять аллели с потерей функции.
Анализ экспрессии ids1 в ушах мутанта ids1 – mum .( A ) РНК-гель, содержащий 1 мкг поли (A) + РНК, выделенную из 2-сантиметровых ушей из A632 ( 1 ), ids1 – mum2 ( 2 ) и ids1 – mum1 ( 3 ) подвергали блоттингу и гибридизовали с 3′-частью кДНК ids1 . ( B ) Контрольная гибридизация. Блот в A зондировали кДНК убиквитина кукурузы, чтобы показать относительную нагрузку.
Для более точной локализации временного и пространственного паттерна экспрессии ids1 мы использовали гибридизацию in situ.На рисунке А показаны продольные срезы вегетативного побега дикого типа.
apex зондировали меченной дигоксигенином антисмысловой РНК, полученной с 3′-конца клона ids1 . Положение листьев внутри побега кукурузы было описано с помощью индекса пластохрона (Lamoreaux et al. 1978), в котором самый молодой лист обозначен как пластохрон 1 или P 1 , а последующие листья — как P 2 , P 3 и т. Д. Положение зарождающегося листа в меристеме обозначается как P 0 . ids1 экспрессия обнаруживается на флангах меристемы в дискретной зоне (рис. A). С позиции самого молодого листа
(обозначено как P 1 ), мы предполагаем, что эта зона экспрессии меристемы находится в положении, в котором будет инициироваться следующий лист (обозначенный как P 0 ). Выражение также наблюдается на кончиках следующих двух более старых листьев, P 1 и P 2 .
Локализация in situ в вегетативных и цветочных верхушках B73 с использованием 3′-части кДНК ids1 .( A ) Апикальная меристема побега с молодыми зачатками листьев (P 1 и P 2 ) и зарождающимися зачатками листьев (P 0 ). Пруток, 65 мкм. ( B ) Молодое соцветие початка. ids1 экспрессия может быть замечена в акропетально инициирующих зачатках пары колосков. Пруток, 70 мкм. ( C ) Колосковые неразветвленные меристемы. Выражение можно увидеть в меристеме колосков, но не в чешуях. Пруток, 36 мкм. ( D ) Ветвящиеся меристемы колосков. Сильное выражение можно увидеть во внутренней и внешней леммах, а также в зоне между
инициирующие верхние и нижние соцветия (стрелка без надписи).Эта зона экспрессии постепенно исчезает в более старом колоске.
ниже. В инициирующих цветочных меристемах экспрессии не обнаружено. Пруток, 70 мкм. ( E ) Цветочек верхний с зарождающимися тычинками. Дискретные области экспрессии ids1 могут наблюдаться в инициирующих зачатках тычинок и палеа. ids1 Выражение сохраняется в расширяющихся внутренних и внешних леммах. Пруток, 70 мкм. ( F ) Старший цветочек с лодикулами. Выражение тычинок теперь локализуется в камерах локул пыльника.Никакого выражения не может быть
замечено в лодикулах (ло). Пруток, 70 мкм. ( G ) ids1 – mum2 колоск, зондированный с ids1. Мало или не может быть обнаружено ids1 расшифровки. Стрелки указывают на соцветия. Пруток, 130 мкм.
В продольных срезах ткани цветка экспрессия ids1 обнаруживается в нескольких различных зачатках боковых органов, а также в паре колосков и меристемах колосков.На рисунке B продольные срезы меристем ранних соцветий, исследованные с помощью ids1 , показывают полосы экспрессии в повторяющемся паттерне, который, очевидно, соответствует зачаткам пары колосков. На более позднем этапе
после образования ножек и сидячих колосков экспрессия ids1 обнаруживается по всей меристеме колосков, но не в первых двух зачатках боковых органов, образованных из колоска.
меристема, внешняя и внутренняя чешуйки (рис.C). Вскоре после зарождения цветочных меристем (рис. D) экспрессия ids1 присутствует в инициирующих внутреннем и внешнем листьях прицветника леммы, прилегающих к цветкам, но отсутствует в
Сама цветочная меристема. Кроме того, видна сильная полоса экспрессии в области между верхними и нижними цветочками.
(Рис. D, стрелка без надписи). У немного более старых колосков, как показано в нижней части рисунка D, эта полоса экспрессии начинается
исчезнуть.Микрофотография колосков на эквивалентных стадиях, полученная с помощью сканирующего электронного микроскопа.
соцветия (рис. E), дискретные группы клеток меристемы цветков, которые образуют тычинки, а палеа экспрессирует
ids1. Экспрессия сохраняется на более поздних стадиях в более старых расширенных палеа и лемма, а также в камерах локул пыльника (рис.
F). В лодикулах или гинеце уха экспрессия была незначительной или отсутствовала (рис.F; данные не показаны). Как показано на рисунке G,
когда зонд anti-sense ids1 используется на колосках ids1 – mum , экспрессия ids1 не обнаруживается.
Обсуждение
Регулирование количества цветков, инициированных меристемой колосков, является важным шагом в определении морфологии колосков.Мутации в гене ids1 влияют на детерминированность меристемы колосков. Вместо того, чтобы производить только верхний и нижний соцветия, меристема колосков ids1 – mum продолжает инициировать дополнительные соцветия в дистихической филлотаксии. Понимание того, как ids1 способствует определению меристемы колосков, требует изучения различных моделей инициации цветков кукурузы.
Одна модель зарождения цветков предполагает, что нижний цветочек начинается латерально, тогда как верхний цветочек — терминальный,
в результате превращения меристемы колосков в верхний цветочек (рис.A) (ирландский язык, 1997 г.). У некоторых видов трав, таких как anthoxanthum, , колоск заканчивается цветком, а семяпочка является конечной структурой (Clifford 1987). Если верхний цветочек является терминальной структурой в нормальном развитии кукурузы, белок IDS1 может действовать, способствуя трансформации.
меристемы колосков в верхний цветочек. IDS1 может делать это самостоятельно или посредством активации идентичности цветочной меристемы.
гены, такие как ортологи LEAFY и APETELA1 , которые могут преобразовывать меристемы соцветий в цветочные меристемы (Mandel and Yanofsky 1995; Weigel and Nilsson 1995).Если бы это было так, мы бы предположили, что трансформация меристемы колосков в меристему цветков задерживается у мутантов ids1 – mum , позволяя меристеме колосков сохраняться и инициировать больше цветков. Такое толкование использовалось
объяснить фенотип доминантных мутантов кукурузы Ts6 , у которых образуются лишние соцветия (Irish 1997). Мутанты Ts6 отличаются от мутантов ids1 – mum , однако, тем, что они также влияют на определение пола и вызывают отсутствие подавления карпеля внутри кисточки,
но не в ухо.
Модели для инициации цветков кукурузы. ( A ) Конечная модель верхнего цветочка. Колосковая меристема разветвляется один раз латерально, образуя нижний цветочек. Остаточный колоск
меристема (показанная черным) затем трансформируется в верхний цветочек (черный). ( B ) Модель бокового разветвления.Меристема колосков имеет боковое ветвление, образуя нижние, а затем верхние соцветия. В
остаточная меристема колосков (показана черным) находится в небольшой области между цветками. Зачаток колоска
меристема находится в рахилле в зрелом колоске (черный).
Альтернативная модель предполагает, что и верхние, и нижние соцветия являются латеральными продуктами меристемы колосков.Два
соцветия кукурузы интерпретировались как боковые ветви рахиллы с небольшим остатком оси или без остатка оси между ними
цветочки (Bonnett 1953). Если ветвление верхнего цветочка латеральное, то в рахилле может быть обнаружен сильно редуцированный зачаток меристемы колосков.
после зарождения верхнего цветочка (рис. Б). Согласно этой модели IDS1 будет участвовать в подавлении неопределенного роста.
внутри меристемы колосков, что препятствует регенерации меристемы колосков после ветвления верхнего цветочка.У мутантов ids – mum остаточная меристема колосков пролиферирует и продолжает образовывать соцветия.
На основании паттерна экспрессии ids1 и времени появления дефектов ids1 – mum , мы отдаем предпочтение второй модели и предполагаем, что функция IDS1 подавляет неопределенный рост в колоске.
меристема. Согласно первой модели, переключение судьбы меристемы колосков происходит только после инициирования нижнего цветочка.Однако дефекты в меристемах колосков ids1 – mum впервые наблюдались в меристемах колосков до зарождения нижних цветков (Рис. E). Сроки
дефект указывает на то, что изменение в детерминации судьбы происходит раньше, в соответствии с экспрессией ids1 , наблюдаемой в меристеме колосков до того, как происходит ветвление цветков (Fig. C). Первая модель также предполагает, что
вся меристема колосков превращается в верхний цветочек, не оставляя остаточной меристемы колосков.Тем не менее, экспрессия ids1 была обнаружена в зоне между верхними и нижними соцветиями у дикого типа (рис. D), в которой остаточный колоск
ожидается, что меристема будет расположена. Дополнительное свидетельство того, что эта область представляет собой остаточную меристему колосков, появляется.
из наблюдения, что он регенерирует после ветвления цветков у мутантов ids1 – mum (Рис. F, G). Более того, ожидание от первой модели состоит в том, что ids1 будут выражены по всему будущему верхнему цветку, чтобы способствовать его преобразованию в цветочную идентичность.Вместо этого, как показано
на рисунке D мы не видим экспрессии ids1 ни в верхней, ни в нижней цветочной меристеме. Наконец, если детерминированность меристемы колосков просто
задержка у мутантов ids1 – mum в соответствии с первой моделью, можно было бы ожидать, что рахилла в конечном итоге оканчивается цветком. Это было
не наблюдается ни у самца, ни у самки ids1 – мама колосков (рис. F; данные не показаны).
Считается, что развитие критического размера меристемы у кукурузы дает сигнал к возникновению ветвления (Sundberg and Orr 1996). Одним из механизмов, с помощью которого IDS1 может подавлять неопределенный рост, является ограничение размера остаточной меристемы колосков.
такой, что он больше не может начинать соцветия. У мутантов ids1 – mum размер остаточной меристемы колосков может быть нарушен и увеличиваться до большего, чем обычно.Колоск большего размера
меристема потенциально может позволить большему количеству событий ветвления произвести дополнительные соцветия. В поддержку этой идеи
ids1 – mum меристема колосков кажется более удлиненной, чем обычно, до образования цветков (Рис. E). Также более крупная меристема колосков
диаметр наблюдается вскоре после зарождения первого цветочка у мутантов ids1 – mum (Рис. D).
Однако важно понимать, что судьба меристемы не всегда зависит от размера меристемы.Например, верхняя
Цветочная меристема намного больше, чем нижняя цветочная меристема во время инициации (Рис. B), и тем не менее обе они одинаково
решил сделать цветочки. Таким образом, наличие более крупной меристемы колосков не обязательно наделяет ее большей неопределенностью.
Не менее важным компонентом различия между определенными и неопределенными судьбами меристемы колосков является способность
регенерировать после зарождения цветков.Меристемы с неопределенной судьбой могут проявлять большую способность к самовосстановлению.
независимо от их первоначального размера. Измененная морфология, наблюдаемая в меристемах колосков у мутантов ids1 – mum , может быть просто отражением этой способности.
Возможно, что одним из способов, которым IDS1 функционирует для поддержания детерминированности меристемы колосков, является подавление этих необходимых факторов.
для сохранения неопределенности.Гомеобокс kn1 ген экспрессируется в нескольких типах неопределенных меристематических тканей, но не в определенных органах, таких как листья.
(Смит и др., 1992; Джексон и др., 1994). Мутанты с потерей функции kn1 кукурузы демонстрируют пониженное ветвление метелки и меньшее количество колосков. Постулировалось, что этот фенотип является результатом потери неопределенного
клетки соцветия, которые в отсутствие kn1, приняли определенные судьбы (Kerstetter et al.1997). Если kn1 на самом деле поддерживает неопределенность меристемы, то экспрессия kn1 должна сохраняться в пределах меристемы колосков ids1-mum и демонстрировать расширенный домен. Хотя это наблюдалось (Fig. D), важно отметить, что kn1 обычно экспрессируется как в колосковой, так и в цветочной меристеме (Fig. A, B). Таким образом, тот факт, что домен экспрессии kn1 расширяется у мутантов ids1 – mum , может просто отражать дополнительные области инициации цветков.
Ген ids1 был клонирован по гомологии с геном AP2 из Arabidopsis и, как было показано, содержит два повтора высококонсервативного домена AP2. Хотя белок IDS1 показывает большее сходство с
AP2 по сравнению со всеми другими AP2 -подобными генами, клонированными до сих пор, ген ids1 может не представлять фактический ортолог AP2 кукурузы.Гомология аминокислот между IDS1 и AP2 не распространяется за пределы домена AP2. Кроме того,
Экспрессия ids1 не была обнаружена в плодолистиках, как обнаружено для AP2 (Jofuku et al. 1994). Наконец, предварительные эксперименты показывают, что сверхэкспрессия кДНК ids1 в Arabidopsis не дополняет мутантный фенотип ap2-1 (G. Chuck unpubl.). Дополнительные члены семейства AP2 , вероятно, будут присутствовать в геноме кукурузы, потому что гибридизации с низкой строгостью с доменом ids1 AP2 в качестве зонда на саузерн-блотах кукурузы показывают несколько полос гибридизации (данные не показаны).Кроме того, несколько выраженных
Было показано, что теги последовательностей из библиотек кДНК кукурузы содержат домены AP2 (Klucher et al. 1996).
Хотя ids1 экспрессируется в зачатках вегетативных и цветочных боковых органов, явных мутантных фенотипов в этих органах обнаружено не было.
В этих органах может действовать определенный уровень генетической избыточности, который может быть обнаружен только путем анализа двойного
мутанты.Например, двойные мутанты ap2 и aintegumenta демонстрируют фенотипы цветочных органов, не наблюдаемые ни у одного единственного мутанта в Arabidopsis (Elliot et al. 1996). Учитывая большое количество AP2 -подобных генов, присутствующих в геноме Arabidopsis (Okamuro et al. 1997), неудивительно, что такая функциональная избыточность встречается у кукурузы. Недавно было высказано предположение, что
два тесно связанных гена MADS-бокса, zag1, и zmm2, , играют избыточную роль в определении идентичности плодолистиков и тычинок кукурузы (Mena et al.1996).
Хотя органы цветков ids1 – мама соцветий кисточки полностью функциональны и напоминают цветки дикого типа, дефекты органов цветков наблюдались у ids1 – мама женских колосков. Возможное объяснение этого дефекта может заключаться в том, что у нормальной самки происходит выкидыш нижнего цветочка.
колоски. Если различие между верхними и нижними цветочками утрачено у мутантов ids1 – mum , то все цветочки могут получить сигнал об прерывании беременности, который обычно присутствует только в нижних цветках дикого типа.Для проверки этой модели будет полезен анализ двойных мутантов с мутантом с метелкой 2 , у которого не происходит выкидыша нижних цветков (DeLong et al. 1993).
В свете того факта, что многие представители семейства трав обладают неопределенными колосками, есть соблазн предположить, что AP2 -подобные гены играют роль в регуляции количества цветков у других трав. Колоски с множеством цветков считаются древним
признак, учитывая, что производные виды почти повсеместно эволюционировали, уменьшив количество цветков (Стеббинс, 1987).Возможный механизм такого снижения может включать изменение функции или домена экспрессии гена ids1 , чтобы включить меристему колосков в дополнение к различным латеральным органам, где он обычно экспрессируется. Анализ
экспрессии ids1 как в современных, так и в древних травах будет интересно проанализировать в этом отношении.
Материалы и методы
Клонирование гена
ids1
Сорок тысяч бляшек библиотеки кДНК B73, построенных из ушей соцветий размером от 1 до 3 см (Jackson et al.1994) были исследованы при пониженной строгости при 55 ° C в 50% формамиде (Schmidt et al. 1993) с полноразмерной кДНК AP2 из Arabidopsis (Jofuku et al. 1994). Было выделено примерно 75 положительных клонов. Далее был проанализирован только самый сильный гибридизирующий класс. Представитель
клон из этого класса был использован для исследования библиотеки кДНК вегетативной меристемы B73 (Lambda ZapII, Stratagene), из которой полноразмерный
Была выделена кДНК размером 1,9 т.п.н. КДНК была субклонирована в pSK- и обе цепи секвенировали с помощью секвенатора ABI.Анализ последовательности
было сделано с использованием GCG (Wisconsin Genetics Group) и сервера электронной почты Национального центра биотехнологии.
Гель-блоты РНК и гибридизация in situ
РНК
выделяли из зародышей, ушей, вегетативных меристем и корней, как описано ранее (Kerstetter et al. 1994). Гибридизацию in situ проводили, как описано Jackson et al.(1994) с использованием 3′-концевого зонда между нуклеотидами 1330 и 1835 вне консервативного домена AP2 . Этот же фрагмент ДНК использовали для зондирования всех гель-блотов РНК. Наблюдается единственная полоса, гибридизирующаяся с этим зондом.
на ДНК-гель-блотах (данные не показаны), поэтому перекрестная гибридизация с другими членами семейства AP2 маловероятна.
Выделение рецессивных аллелей
ids1 методом ПЦР
Приблизительно 42000 растений F 1 растений из скрещиваний между запасами, содержащими активные мобильных элементов Mu , были выращены и подвергнуты скринингу с помощью ПЦР в Pioneer Hi-Bred International для вставки в ID1 (Bensen et al.1995) с праймером ZAP2-1 (5′-CCGGTGGCGCCAGCGAAGAA-3 ‘) и Mu-9242 (5′-CCCTGAGCTCTTCGTC (CT) ATAATGGCAATTATCTC-3’), вырожденный
праймер, который связывается с концевым инвертированным повтором Mu. реакций ПЦР проводили на гелях, блотировали и зондировали с помощью кДНК ids1 . Продукты ПЦР, которые гибридизуются с кДНК ids1 , идентифицировали людей, которые, вероятно, имеют вставки Mu в ген ids1 . Такой скрининг идентифицировал девять кандидатов с использованием праймера ZAP2-1.ДНК-гель-блот-анализ расщепленной ДНК, полученной из
самоопыляемое потомство каждого кандидата выявило полиморфизмы только для двух из девяти семейств при зондировании с помощью кДНК ids1 . Эти же два семейства были единственными, которые генерировали ids1 продуктов ПЦР с использованием праймеров ZAP2-1 и Mu 9242. Вероятно, что семь других семейств, которые не показали полиморфизмов
представляют собой события соматической вставки Mu в ids1 , которые не передались в зародышевую линию.Каждый аллель ids1 – mum был получен от скрещивания активной линии Mu и инбредной линии A632. Оба аллеля были однократно скрещены с A632 и подверглись самоопылению для наблюдения фенотипов.
у гомозигот. Фенотипы также наблюдались после второго обратного скрещивания с A632 и обратного скрещивания с инбредным W23.
Сканирующая электронная микроскопия
Ткань фиксировали в FAA (50% этанол, 5% уксусная кислота, 3.7% формальдегида) при 4 ° C в течение ночи и обезвоживание в этаноле
серию до 100%. Затем образцы были высушены до критической точки и напылены палладием. Образцы были просмотрены на ISI
30 РЭМ при ускоряющем напряжении 10 кВ.
Благодарности
Мы благодарим Дж. Окамуро и Д.Джофуку (Калифорнийский университет, Санта-Крус) за дар кДНК AP2 , Р. Керстеттеру за использование его РНК-блоттинга и К. Канаде (Pioneer Hi-Bred International) за идентификацию вставок ids1 . Мы также признательны Э. Фольбрехту, П. Макстину, Л. Райзеру, Ф. Хемпелю и сотрудникам лаборатории Хека.
за рецензирование рукописи и полезные обсуждения. G.C. был поддержан стипендией Калифорнийского университета.
Работа поддержана Институтом У.S. Департамента сельского хозяйства и частично выполняются в рамках Совместных исследований и разработок.
Соглашение с Pioneer Hi-Bred International, Inc.
Расходы на публикацию этой статьи были частично покрыты за счет оплаты страницы. Поэтому эта статья должна быть настоящим
помечены как «реклама» в соответствии с разделом 1734 Кодекса США 18 исключительно для обозначения этого факта.
MULTI-FLORET SPIKELET1, который кодирует белок AP2 / ERF, определяет судьбу колосковой меристемы и идентичность стерильной леммы в рисе
- © Американское общество биологов растений, 2013.Все права защищены.
Абстракция
Колосок — это уникальная структура соцветия травы. Молекулярный механизм, контролирующий развитие колоска, остается неясным. В этом исследовании мы идентифицировали мутант по колоску риса ( Oryza sativa ), multi — цветочек spikelet1 ( mfs1 ), который показал замедленную трансформацию меристем колосков в цветочные меристемы, что привело к появлению дополнительных шелковидных орган и удлиненная рахилла.Кроме того, стерильная лемма гомеотически преобразовалась в рудиментарную чешуйку, а тело палеа дегенерировало в mfs1 . Эти результаты предполагают, что ген MULTI — FLORET SPIKELET1 ( MFS1 ) играет важную роль в регуляции детерминированности меристемы колосков и идентичности органов цветка. MFS1 принадлежит к кладе с неизвестной функцией в семействе APETALA2 / этилен-чувствительный фактор (AP2 / ERF). Слитый белок MFS1-зеленый флуоресцентный белок локализован в ядре. MFS1 матричная РНК экспрессируется в различных тканях, особенно в меристемах колосков и цветков. Более того, наши результаты показывают, что MFS1 положительно регулирует экспрессию LONG STERILE LEMMA и INDETERMINATE SPIKELET1 ( IDS1 ) -подобных генов SUPERNUMERARY BRACT и OsIDS1 .
В репродуктивной фазе покрытосеменных меристема побега трансформируется в меристему соцветия, которая затем дает цветочную меристему, из которой начинают развиваться цветочные органы в соответствии с механизмом, известным как модель ABCDE (Coen and Meyerowitz, 1991; Coen и Nugent, 1994; Dreni et al., 2007; Ohmori et al., 2009). Соцветие можно классифицировать как детерминантное или неопределенное в зависимости от того, трансформирована ли его апикальная меристема в конечную цветочную меристему. В неопределенном соцветии боковая меристема постоянно дифференцируется от апикальной меристемы, которая не превращается в конечную цветочную меристему, как это происходит во время развития соцветий Arabidopsis ( Arabidopsis thaliana ) и львиного зева ( Antirrhinum majus ). .Напротив, в детерминированном соцветии апикальная меристема трансформируется в конечную цветочную меристему после образования фиксированного количества боковых меристем, как это происходит во время развития соцветий табака ( Nicotiana tabacum ) и томатов ( Solanum lycopersicum ; Bradley et al., 1997; Ratcliffe et al., 1999; Sussex and Kerk, 2001; Chuck et al., 2008).
В целом соцветия трав состоят из ветвей и колосков (Coen, Nugent, 1994; Itoh et al., 2005; Кобаяши и др., 2010). У этих организмов меристема ветвей детерминирована. Он дает несколько боковых меристем колосков, за которыми следует конечная меристема колосков. Колосок, специфическая единица соцветия травы, состоит из пары прицветников и от одного до 40 цветков; он показывает определенность или неопределенность в зависимости от вида (Clifford, 1987; Malcomber et al., 2006). У видов с детерминированным колоском, таких как рис ( Oryza sativa ), после образования фиксированных боковых цветочных меристем меристемы колосков превращаются в терминальные цветочные меристемы, что приводит к прекращению судьбы меристемы колосков.Напротив, у видов с неопределенным колоском, таких как пшеница ( Triticum aestivum ), судьба меристемы колосков сохраняется постоянно и дает различное количество боковых цветочных меристем.
Было показано, что у Arabidopsis ген TERMINAL FLOWER1 ( TFL1 ) сохраняет неопределенность в судьбе соцветия. У мутанта tfl1 меристемы соцветий были преобразованы в цветочные меристемы раньше, чем у дикого типа, но эктопическая экспрессия TFL1 привела к трансформации цветочных меристем на более поздней стадии развития во вторичные меристемы соцветий (Bradley et al. al., 1997; Рэтклифф и др., 1999; Mimida et al., 2001). У риса сверхэкспрессия одного из -подобных генов TFL1, RICE CENTRORADIALIS1 ( RCN1 ) или RCN2 задерживает переход меристем ветвей в меристемы колосков и, наконец, приводит к образованию большего количества ветвей. и колоски, чем у дикого типа (Nakagawa et al., 2002; Rao et al., 2008).
На сегодняшний день не зарегистрировано ни одного гена, который поддерживает неопределенность меристемы колосков.Однако было показано, что два класса генов участвуют в прекращении неопределенности меристем колосков. Один из них — группа генов идентичности терминальных меристем цветков. Специфическая для травы кладка LEAFY HULL STERILE1 ( LHS1 ) из подсемейства SEPALLATA ( SEP ) относится к этому классу. LHS1 -подобные гены, как было обнаружено, экспрессируются только в терминальной цветочной меристеме у видов с детерминированностью колосков, что предполагает, что они исключительно определяют продукцию терминальной цветочной меристемы, посредством которой колосковая меристема приобретает детерминированность (Cacharroón et al., 1999; Малкомбер и Келлог, 2004 г .; Зан и др., 2005). Другой класс включает гены, подобные INDETERMINATE SPIKELET1 ( IDS1 ), которые принадлежат к семейству APETALA2 / этилен-чувствительный фактор ( AP2 / ERF ). В отличие от LHS1 -подобных генов, этот класс генов регулирует правильное время перехода меристемы колосков в меристему цветков, но не определяет идентичность терминальной меристемы цветков. У кукурузы ( Zea mays ) потеря функции IDS1 дает дополнительные соцветия (Chuck et al., 1998). Кроме того, мутация SISTER OF IDS1 ( SID1 ), паралога IDS1 кукурузы, не привела к дефектам развития колосков. Однако двойной мутант ids1 + sid1 неспособен генерировать органы цветков и вместо этого развивает более прицветниковые структуры, чем обнаруживаются у растений дикого типа (Chuck et al., 2008). Геном риса содержит два IDS1 -подобных гена, SUPERNUMERARY BRACT ( SNB ) и OsIDS1 .Потеря активности SNB или OsIDS1 приводила к образованию дополнительных рудиментарных чешуек, а у snb + osids1 двойных мутантных растений развивались более рудиментарные чешуи, чем у любого из его родительских мутантов (Lee et al., 2007; Lee and An, 2012). Эти результаты показали, что мутировавшие IDS1 -подобные гены продлевали активность меристемы колосков.
У большинства представителей Oryzeae колосок отличается от колосов других злаков тем, что он состоит из пары рудиментарных чешуек, пары стерильных чешуек (пустых чешуек) и одного цветочка (Schmidt and Ambrose, 1998; Ambrose et al. al., 2000; Келлог, 2009; Hong et al., 2010). Рудиментарные чешуйки обычно считаются сильно редуцированными прицветниками, но происхождение стерильных чешуек широко обсуждается. Недавние исследования показали, что стерильные леммы являются рудиментарными леммами двух боковых цветочков. Ген LONG STERILE LEMMA ( G1 ) / ELONGATED EMPTY GLUME1 ( ELE1 ) является членом семейства генов, специфичных для растений. У мутанта g1 / ele1 стерильные леммы гомеотически трансформировались в леммы (Yoshida et al., 2009; Hong et al., 2010). Было показано, что гены OsMADS34 и EXTRA GLUME1 ( EG1 ) определяют идентичность стерильных лемм. У мутантов osmads34 и eg1 стерильные леммы были увеличены и приобрели идентичность лемм (Li et al., 2009; Gao et al., 2010; Kobayashi et al., 2010). Кроме того, SEP -подобный ген LHS1 / OsMADS1 , который определяет идентичность как леммы, так и палеи, не выражался в стерильных леммах, а эктопическая экспрессия в стерильных леммах приводила к трансформации стерильных лемм в леммы (Jeon et al., 2000; Ли и др., 2009; Танака и др., 2012). Эти результаты позволяют предположить, что лемма о стерильности может быть гомологичной лемме. Тем не менее, некоторые исследователи все еще считали, что стерильные леммы представляют собой рудиментарные прицветники, похожие на рудиментарные чешуйки (Schmidt and Ambrose, 1998; Kellogg, 2009; Hong et al., 2010).
В этом исследовании мы выделили ген риса MULTI — FLORET SPIKELET1 ( MFS1 ), который принадлежит к кладе с неизвестной функцией в семействе генов AP2 / ERF .Было показано, что мутация MFS1 задерживает трансформацию меристемы колосков в меристему цветков и приводит к дегенерации стерильной леммы и палеи. Эти результаты подтверждают, что MFS1 играет важную роль в регуляции детерминированности колосков и идентичности органов. Наши результаты также показывают, что MFS1 положительно регулирует экспрессию G1 и IDS1 -подобных генов SNB и OsIDS1 .
РЕЗУЛЬТАТЫ
Мы идентифицировали два рецессивных мутанта, связанных с развитием рисового колоска, mfs1-1 и mfs1-2 (Рис. 1; Дополнительный Рис. S1). Тест на аллелизм показал, что два мутанта были аллельными. Учитывая, что mfs1-1 обнаружил более серьезные дефекты, чем mfs1-2 , остальная часть этой статьи сосредоточена в первую очередь на мутанте mfs1-1 .
Рисунок 1.
Фенотипы колосков у дикого типа и mfs1-1 .A и B, Колосок дикого типа. C-1, Эпидермальная поверхность палеи дикого типа. C-2, Эпидермальная поверхность леммы дикого типа. D и E, Гистологический анализ колосков дикого типа. F и G, mfs1-1 колоск с дегенерированной палеей. H, Эпидермальная поверхность дегенерированного палеа у G. I, mfs1-1 колоск с дополнительной оболочкой и нормальным палеа. J. Гистологический анализ колоска mfs1-1 с дополнительной оболочкой и нормальной палеей. K and L, mfs1-1 колоск с дополнительной оболочкой и вырожденными бледными краями.M, Эпидермальная поверхность дегенерированной палеа в L. N и O, гистологический анализ колоса mfs1-1 с дополнительной оболочкой и дегенерированными палеями. P, Эпидермальная поверхность стерильной леммы и рудиментарной чешуи у дикого типа. От Q до T, эпидермальная поверхность дегенерированной стерильной леммы и рудиментарной чешуи в mfs1-1 . U, относительные уровни экспрессии генов идентичности органов цветка в органах цветка дикого типа (wt) и mfs1-1 . dp, дегенерированная палеа; dsl, вырожденная стерильная лемма; эр, удлиненная рахилла; hl, корпус (lemma / palea) -подобный орган; ле, лемма; вот, lodicule; па, палеа; приятель, палеоподобный орган; пи, пестик; rg, рудиментарная чешуйка; sl, стерильная лемма; ул, тычинка.Черные стрелки обозначают сосудистые пучки. Полосы = 1000 мкм в A, B, F, G, I, K и L и 100 мкм в C – E, H, J, M и N – T. Столбцы ошибок в U указывают на SD.
mfs1-1 Демонстрирует плейотропные дефекты в развитии колосков
Обычно рисовые колоски дикого типа состоят из одной пары рудиментарных чешуек, одной пары стерильных чешуек и одного конечного плодородного цветочка. Цветочек состоит из четырех оборотов цветковых органов: одной леммы и одной палеи в обороте 1, двух лодикул в обороте 2, шести тычинок в обороте 3 и одного пестика с двумя рыльцами в обороте 4 (рис.1, А и Б).
У колосков дикого типа стерильная лемма была больше, чем рудиментарная чешуйка (рис. 1А). Большая часть эпидермиса стерильной леммы была гладкой и состояла из регулярно расположенных плоских клеток и редких клеток с трихомами на абаксиальной стороне (рис. 1P). Эпидермальные клетки рудиментарных чешуек расположены нерегулярно и несут множество выступов и трихом (рис. 1P). Напротив, стерильная лемма была уменьшена в разной степени, даже напоминая по размеру рудиментарную чешуйку у мутанта mfs1-1 (рис.1, F, I и L; Дополнительный рис. S1). Обильные выпячивания и трихомы находились на среднем и нижнем эпидермисе дегенерированных стерильных чешуек, которые были очень похожи на таковые из рудиментарной чешуи (рис. 1, Q – T). Между тем, регулярные и гладкие клетки, как и клетки стерильной леммы дикого типа, по-прежнему оставались в верхней части вырожденных стерильных лемм (рис. 1, Q – T). Эти результаты показали, что вырожденная стерильная лемма у мутанта mfs1-1 имела идентичность как стерильных лемм, так и рудиментарных чешуек.
Было обнаружено, что 65% из mfs1-1 колосков развили дополнительный корпус (lemma / palea) -подобный орган (рис. 1, I и K; дополнительный рис. S1A). Лемма дикого типа имела четыре клеточных слоя, силикатные клетки, фиброзную склеренхиму, губчатые паренхиматозные клетки и несиликатные клетки, а также развивала пять сосудистых пучков. По сравнению с леммой, палеа имела три сосудистых пучка и состояла из двух частей: тела палеа (bop) и двух краевых областей палеа (mrp). Клеточная структура bop была очень похожа на структуру леммы, но mrp демонстрировала характерный гладкий эпидермис, в котором отсутствовало эпикутикулярное силикатизированное утолщение, обнаруженное в lemma и bop (рис.1, Б – Д). У мутанта mfs1-1 дополнительный корпусообразный орган обнаруживал сходную гистологическую структуру и имел пять сосудистых пучков, напоминающих лемму дикого типа (рис. 1, J и N). Мы обнаружили уровни мРНК гена идентичности lemma DROOPING LEAF ( DL ), генов идентичности lemma и palea OsMADS1 , OsMADS14 и OsMADS15 и гена идентичности mrp mrp OsMAD. -1 дополнительных корпусных органов.Обнаружены избыточные уровни транскриптов OsMADS1 , OsMADS14 , OsMADS15 и DL , но не было обнаружено экспрессии OsMADS6 (рис. 1U). Эти результаты показали, что дополнительный корпусообразный орган имел тождество леммы.
Из mfs1-1 колосков с дополнительными леммоподобными органами, 38% не имели нормальной палеа. Два палеоподобных органа наблюдались в положении, обычно занимаемом палеей (рис.1, K и L; дополнительный рис.S1A). Интересно, что каждый палеоподобный орган состоял из двух mrps и меньшего bop с двумя сосудистыми пучками (Fig. 1, L – O). Каждый mrp и bop имел текстуру, аналогичную структуре палеи дикого типа (рис. 1, M и N). OsMADS1 , OsMADS14 и OsMADS15 экспрессировались нормально (рис. 1U), тогда как DL не экспрессировался в палеоподобных органах, как и в случае с палеа дикого типа (рис. 1U). . OsMADS6 экспрессия была более интенсивной в палеоподобных органах mfs1-1 , чем в палеоподобных органах дикого типа (рис.1U). Эти находки указывают на то, что палеоподобные органы были дегенерированными бледными, и повышенная экспрессия OsMADS6 в палеоподобных органах, вероятно, была вызвана относительным обилием тканей mrp.
У 21% из mfs1-1 колосков палея была дегенерирована в различной степени (дополнительный рис. S1A). Большая часть дегенерированных палео содержала нормальный mrp и восстановленный bop (дополнительный рис. S1, D и E). В нескольких случаях дегенерированная палеа сохраняла только mrp-подобные структуры, которые содержали несиликатный верхний эпидермис без трихом и выступов (рис.1, F – H). Эти результаты позволяют предположить, что развитие mfs1-1 bop серьезно пострадало.
Одновременно мы исследовали дефекты трех внутренних оборотов у мутанта mfs1-1 . У цветков (41%) с нормальными цветками идентичность и количество органов трех внутренних оборотов не изменились (дополнительная таблица S2). У цветков (59%) с дегенерированными цветками количество органов трех внутренних оборотов было различным, но они сохранили свою идентичность (дополнительная таблица S2; дополнительный рис.S1, F – I). Кроме того, 81% из колосков mfs1-1 имели удлиненные рахиллы (рис. 1, F, I, Q и S; дополнительный рис. S1A).
mfs1-1 Показано аномальное раннее развитие колосков
Мы исследовали молодые колоски дикого типа и мутантов на разных стадиях развития с помощью сканирующей электронной микроскопии. На стадии 4-го колоска (Sp4) лемма и зачатки палеа у цветка дикого типа начали развиваться, а верхушка леммы была выпуклой и была крупнее палеа (рис.2А). У мутанта mfs1-1 некоторые колоски развили лишние леммоподобные зачатки органов, и их палеэ были либо нормальными (Fig. 2I), либо дегенерированными (Fig. 2M). В то же время цветочная меристема была увеличена на участках колосков с дегенерированными бледными краями (рис. 2М). У других колосков не было лишнего леммоподобного органа, тогда как их палеа зачатки были уменьшены в размерах (рис. 2E). Во время Sp5 и Sp6 цветок дикого типа образовал шесть сферических зачатков тычинок; Развитие одного зачатка тычинок на стороне леммы задерживалось, тогда как других развивалось синхронно (рис.2Б). Не наблюдалось значительных различий у цветков с лишним леммоподобным органом и нормальной палеа (рис. 2J). Однако у некоторых цветков с дополнительными леммоподобными органами и аномальными пледами развитие тычинок не было синхронным, и количество тычинок варьировало (рис. 2, N и O; дополнительный рис. S1, F и G; дополнительная таблица S2). У цветков с дегенерированными бледными пятнами мы не обнаружили явных дефектов, за исключением количества тычинок (рис. 2F; дополнительная таблица S2). На стадиях Sp7 и Sp8 (формирование зачатков пестика) лемма и палеа перешли в дальнейшую стадию развития.У мутанта mfs1-1 наблюдались очевидные лишние леммоподобные органы и дегенерированные палеэ (рис. 2, H, L и P).
Рисунок 2.
колосков на ранних стадиях развития у дикого типа и mfs1-1 . От A до D, колоски дикого типа на стадиях Sp4 (A), от Sp5 до Sp6 (B), Sp7 (C) и Sp8 (D). От E до P, mfs1-1 колоск на стадиях Sp4 (E, I и M), от Sp5 до Sp6 (F, J и N), Sp7 (G, K и O) и Sp8 (H, L , и P). dsl, Вырожденная стерильная лемма; эл — дополнительный орган, похожий на лемму; fm — цветочная меристема; ле, лемма; вот, lodicule; па, палеа; приятель, палеоподобный орган; пи, пестик; sl, стерильная лемма.Звездочками обозначены тычинки. Штанги = 100 мкм.
Мы также исследовали стерильную лемму на разных стадиях развития. На стадиях от Sp4 до Sp6 стерильная лемма была крупнее рудиментарной чешуи у дикого типа (рис. 3, A и B). У мутанта mfs1-1 размер стерильной леммы был подобен размеру рудиментарной чешуи (рис. 3, E и F). На стадиях Sp7 и Sp8 стерильная лемма резко дифференцировалась и была намного крупнее рудиментарной чешуи у дикого типа (рис.3, В и Г). Однако стерильная лемма mfs1-1 была меньше, чем у дикого типа, напоминая рудиментарную чешуйку на этих стадиях (Sp7 и Sp8; рис. 3, G и H). Между тем, эпидермальные клетки начали удлиняться в стерильных чешуях и все еще сохраняли свой размер в рудиментарной чешуе у дикого типа (рис. 3D), тогда как их размеры сохранялись как в рудиментарной чешуе, так и в стерильной чешуе mfs1-1. Мутант (фиг. 3H). Эти результаты предполагают, что идентичность стерильной леммы mfs1-1 была затронута, а стерильная лемма у мутанта mfs1-1 обнаружила паттерн развития, подобный таковому у рудиментарной чешуи.
Рисунок 3.
Развитие стерильной леммы у дикого типа и mfs1-1 колосков на ранних стадиях. От A до D, развитие леммы о стерильности у дикого типа на стадиях от Sp4 (A), от Sp5 до Sp6 (B), Sp7 (C) и Sp8 (D). От E до H, развитие стерильной леммы в mfs1-1 на стадиях Sp4 (E), Sp5 до Sp6 (F), Sp7 (G) и Sp8 (H). dsl, Вырожденная стерильная лемма; hl, корпусный орган; ле, лемма; па, палеа; приятель, палеоподобный орган; rg, рудиментарная чешуйка; sl, стерильная лемма. Штанги = 100 мкм.
Паттерны экспрессии генов идентичности цветочных органов на ранних стадиях развития цветка
Паттерны экспрессии DL , OsMADS1 и OsMADS6 , которые, как известно, участвуют в регуляции идентичности lemma и palea, были исследованы на ранних стадиях развития цветка.
В цветках дикого типа DL сначала экспрессировался в зачатках леммы на стадиях от Sp4 до Sp6 (рис. 4, A и B), а затем также в зачатках пестика после SP7 (рис. 4C), тогда как DL транскрипты все еще сохранялись в лемме на Sp8 (рис. 4D). В цветках mfs1-1 сигналы DL были выражены в дополнительном леммоподобном органе в дополнение к лемме и пестику (рис. 4, E – H). Эти данные доказали, что у колосков действительно развивались дополнительные листочки на ранней стадии развития цветка.
Рисунок 4. Экспрессия
гена DL в цветках дикого типа и mfs1-1 . От A до D, цветы дикого типа. От E до H, mfs1-1 цветков. В столбцах 1–3 показаны продольные сечения цветков на стадиях от Sp4 до Sp7, а в столбце 4 показаны поперечные срезы цветков на стадии Sp8. el, лишний леммоподобный орган; ле, лемма; па, палеа; приятель, палеоподобный орган; пи, пестик; vb, сосудистый пучок. Штанги = 50 мкм.
На стадиях от Sp4 до Sp8 OsMADS1 экспрессировался в нижних конечностях и палеоцветах дикого типа (рис.5, А – Г). У мутанта mfs1-1 , OsMADS1 сигналы наблюдались в дополнительном леммоподобном органе, лемме и палеа (Fig. 5, E – H). На стадиях от Sp4 до Sp7 экспрессия OsMADS6 не показала значительных различий между mfs1-1 и цветками дикого типа и была обнаружена в меристеме цветков и зачатках mrp, lodicule и пестика (рис. 5, I – K). и М – О). В Sp8 в поперечном срезе цветков дикого типа транскриптов OsMADS6 были обнаружены в mrp, lodicule и пестике (рис.5L). У мутанта mfs1-1 OsMADS6 сигналов были обнаружены в двух mrps каждого палеоподобного органа, lodicule и пестика (Fig. 5P). Эти результаты также предполагают, что палеоподобные органы произошли от paleae у мутанта mfs1-1 .
Фигура 5.
Экспрессия OsMADS1 и OsMADS6 в цветках дикого типа и mfs1-1 . A – D: экспрессия OsMADS1 в цветках дикого типа. E-H, экспрессия OsMADS1 в цветках mfs1-1 .Экспрессия от I до L OsMADS6 в цветках дикого типа. От M до P, экспрессия OsMADS6 в цветках mfs1-1 . В столбцах 1–3 показаны продольные сечения цветков на стадиях от Sp4 до Sp7, а в столбце 4 показаны поперечные срезы цветков на стадии Sp8. el, лишний леммоподобный орган; fm — цветочная меристема; ле, лемма; вот, lodicule; па, палеа; приятель, палеоподобный орган; пи, пестик. Штанги = 50 мкм.
Молекулярное клонирование и идентификация
MFS1
Ген MFS1 ранее был картирован в области около 350 т.п.н. на хромосоме 5 (Ren et al., 2012). Здесь расположение MFS1 было сужено до физического расстояния 67 т.п.н. между маркерами вставки / делеции IND17 и IND24 (рис. 6А), в которых есть 16 аннотированных генов (http://www.gramene.org /). Анализ секвенирования идентифицировал однонуклеотидную замену от C на T в пределах предсказанного транскрипционного фактора AP2 / ERF ( LOC_Os05g041760 ) в разных положениях двух аллелей mfs1 , вызывая аминокислотные мутации Ala-66 в Val-66. в мутанте mfs1-1 и от Thr-51 до Ile-51 в мутанте mfs1-2 (рис.6А). Чтобы проверить, были ли эти мутации причинно связаны с мутантным фенотипом, был трансформирован геномный фрагмент Os05g041760 дикого типа, который содержал кодирующую последовательность, 2925 п.н. последовательности перед стартовым кодоном и 938 п.н. последовательности ниже стоп-кодона. в мфс1-1 . В результате этого мутантные фенотипы были полностью спасены в трансгенных растениях (дополнительный рис. S2, A – C). Далее мы выполнили РНК-интерференцию (РНКи), чтобы заглушить MFS1 в сорте japonica Zh21.В трансгенных растениях уровень транскрипта MFS1 был значительно снижен (фиг. 6F), и наблюдались плейотропные дефекты колосков, подобные дефектам mfs1-1 (фиг. 6, B – E). Взятые вместе, эти результаты подтвердили, что Os05g041760 является геном MFS1 .
Рисунок 6.
Выделение гена MFS1 и субклеточная локализация белка MFS1. A, Отобразите положение локуса MFS1 . Показаны относительные положения клонов бактериальных искусственных хромосом (ВАС).Ниже представлена геномная структура MFS1 . Показаны сайты мутации в mfs1 . Стрелки указывают сайты предсказанных генов в интервале от IND17 до IND24. Б — Фенотип растений Ж21. C к F, анализ РНКи MFS1 и фенотипов трансгенных растений. C – E, фенотипы трансгенных растений с РНКи. Зеленые стрелки указывают на вырожденную стерильную лемму. dp, дегенерированная палеа; эр, удлиненная рахилла; hl, корпус (lemma / palea) -подобный орган; ле, лемма; па, палеа; sl, стерильная лемма.F, Экспрессия MFS1 в трансгенных растениях с РНКи. WT, Дикий тип. G к L, анализ субклеточной локализации белка MFS1 с использованием протопластов риса. G к I, слитый белок GFP. G, изображение управления цифровым изображением. H, изображение в светлом поле. I. Объединенное изображение слитого белка GFP. J к L, MFS1-GFP. J, изображение управления цифровым изображением. K, изображение в светлом поле. L, объединенное изображение слитого белка MFS1-GFP. Полосы = 1000 мкм от B до E и 50 мкм от G до L.
MFS1 Кодирует белок домена ERF
Генное семейство AP2 / ERF специфично для растений и включает четыре подсемейства: AP2, RAV, DREB, и ERF (Sharoni et al., 2011). Филогенетический анализ показал, что MFS1 и его ортологи из мха, голосеменных, двудольных и злаковых составляют MFS1-подобную кладу, тогда как хорошо известные доменные белки ERF FZP и BD1 и их ортологи составляют другую кладу в подсемействе ERF (рис. 7). . Эти результаты свидетельствуют о том, что гены MFS1 и FZP / BD1 разошлись до появления голосеменных и что гены MFS1 отличаются от хорошо известных генов AP2 / ERF .Кроме того, филогенетический анализ также показал, что другие известные гены домена AP2 ( SNB , OsIDS1 и SHAT1 ) имеют отдаленное эволюционное родство с MFS1 -подобным и FZP / BD1 -подобные гены.
Рисунок 7.
Филогенетическое дерево белков MFS1. Филогенетическое дерево было построено с использованием метода максимального правдоподобия, основанного на матричной модели Джонса-Тейлора-Торнтона. Gy / Mo, голосеменные растения и мхи.[См. Онлайн-статью для цветной версии этого рисунка.]
Анализ последовательности показал, что все MFS1-подобные белки содержат высококонсервативный домен ERF, расположенный близко к их N-концу. Между тем, консервативный C-концевой домен был идентифицирован в MFS1-подобных белках трав и двудольных растений, которые имеют общий мотив DLNEPP 185-190 . Уникальный сайт (V 37 ) и мотив (SPWH 132-135 ) также были идентифицированы в MFS1-подобных белках трав (дополнительный рисунок S3).Кроме того, ген MFS1 имеет низкое сходство последовательности с известными генами AP2 / ERF вне домена AP2 / ERF (дополнительный рисунок S3).
Векторы, которые содержали слитый белок MFS1ORF-GFP, слитый белок SL1ORF-GFP и единственный белок GFP, временно экспрессировались в протопластах риса. Белок SL1ORF-GFP действовал как положительный ядерный генный контроль (Xiao et al., 2009). Зеленая флуоресценция была обнаружена в ядрах протопластов риса для слитых белков MFS1ORF-GFP и SL1ORF-GFP (рис.6, J – L; Дополнительный рис. S2, D – F). В клетках, которые экспрессировали только GFP, зеленая флуоресценция наблюдалась равномерно по всей клетке, кроме вакуоли (рис. 6, G – I). Эти результаты предполагают, что MFS1 кодирует ядерный белок, который может действовать как фактор транскрипции.
Паттерны экспрессии
MFS1
Количественный анализ ПЦР с обратной транскрипцией (qPCR) показал, что MFS1 универсально экспрессируется в различных тканях, включая корни, стебли, листья и метелки, с более высокими уровнями в молодых метелках (2 см или меньше), чем в других исследованных тканях (рис.8А). Кроме того, образец экспрессии MFS1 исследовали с помощью гибридизации in situ. Во-первых, MFS1 высоко экспрессируется в меристемах ветвей и колосков (рис. 8, B – D). Затем сильные сигналы наблюдались в местах инициации стерильного зачатка леммы (рис. 8E). Когда сформировались зачатки леммы и палеа, обильные транскриптов MFS1 были обнаружены в лемме, палеа и цветочной меристеме (рис. 8, D, F и G). Впоследствии экспрессия MFS1 была в основном ограничена леммой, палеей, лодикулой и тычинкой (рис.8, G и H). После образования пестика сигналы MFS1 исчезли с леммы и пестика, но остались в лодикуле, тычинке и пестике (рис. 8, I и J).
Рисунок 8.
Шаблон выражения MFS1 . A, экспрессия MFS1 в различных тканях, как обнаружено с помощью кПЦР. R, корень; S, стержень; L, лист. B к J, гибридизация in situ в метелках и цветках дикого типа с использованием антисмыслового зонда MFS1 . bm — меристема ветвей; fm — цветочная меристема; ле, лемма; вот, lodicule; па, палеа; пи, пестик; rg, рудиментарная чешуйка; sl, стерильная лемма; см, колосковая меристема.Звездочками обозначены тычинки. Штанги = 50 мкм.
MFS1 влияет на экспрессию генов, связанных с развитием колосков
Учитывая, что у мутанта mfs1-1 обнаружены дефекты колосков, мы исследовали уровни экспрессии IDS1 -подобных генов SNB и OsIDS1 , которые тесно связаны с переходом и детерминацией меристемы колосков у риса (Lee et al., 2007; Lee and An, 2012). Транскрипты SNB накапливались преимущественно в молодых метелках длиной менее 2 см, и их уровни были ниже у мутанта mfs1-1 , чем у дикого типа (рис.9А). Затем уровни транскриптов SNB резко снизились в метелках длиной более 2 см, и не было обнаружено различий в уровнях экспрессии SNB между метелками дикого типа и mfs1-1 метелками длиной от 2 до 5 см ( Рис. 9A). OsIDS1 транскриптов впервые были обнаружены в молодых метелках менее 0,5 см, и их больше было в метелках дикого типа длиной от 0,5 до 5 см (фиг. 9A). По сравнению с таковой у дикого типа, экспрессия OsIDS1 не показывала очевидных изменений в метелках с длиной менее 0.5 см, тогда как у mfs1-1 метелок длиной от 0,5 до 5 см она резко уменьшилась (рис. 9А). Эти результаты означают, что MFS1 положительно регулирует экспрессию IDS1 -подобных генов SNB и OsIDS1 .
Фигура 9.
Экспрессия SNB , OsIDS1 и G1 в цветках дикого типа и mfs1-1 . A, анализ qPCR SNB , OsIDS1 и G1 в развивающихся метелках дикого типа (wt) и mfs1-1 метелок на разных стадиях.От B до F, экспрессия G1 в цветках дикого типа. G к K, экспрессия G1 в цветках mfs1-1 . le, лемма; вот, lodicule; па, палеа; sl, стерильная лемма; см, колосковая меристема. Штанги = 50 мкм.
Мы использовали кПЦР для изучения экспрессии гена G1 , который, как было показано, участвует в спецификации идентичности стерильной леммы (Yoshida et al., 2009; Hong et al., 2010). У дикого типа высокий уровень экспрессии G1 был обнаружен в метелках короче 2 см, но уровни мРНК были значительно снижены в метелках размером от 2 до 5 см (рис.9А). В мутанте mfs1-1 , G1 обнаружил более низкие уровни экспрессии в молодых метелках короче 5 см (фиг. 9A). Гибридизация in situ показала, что у дикого типа сигналы G1 сильно детектировались в стерильных леммах на стадии дифференцировки и формирования стерильных зачатков лемм, а впоследствии они заметно уменьшались, когда стерильные леммы начинали удлиняться (рис.9, B). –F). Экспрессия G1 была слабой в стерильных зачатках леммы колоска mfs1-1 во время анализируемых стадий (рис.9, G – K), что согласуется с результатами анализа КПЦР. Эти данные свидетельствуют о том, что MFS1 положительно регулирует экспрессию G1 .
ОБСУЖДЕНИЕ
В этом исследовании мы охарактеризовали новый ген домена AP2 / ERF , MFS1 , который участвует в регуляции детерминации меристемы колосков и идентичности органов цветка у риса. MFS1 способствует экспрессии генов SNB , OsIDS1 и G1 , участвующих в развитии колосков.
MFS1 Влияет на детерминированность меристем колосков
У большинства колосков у мутантных растений mfs1-1 развивается дополнительная лемма. Около 27% этих колосков давали нормальные соцветия после появления дополнительных лемм, что свидетельствует о том, что эти колоски состояли из конечного цветочка и дегенерированного бокового цветочка, который содержал только лемму. У других колосков с аномальными цветками наблюдались два палеоподобных органа, что соответствовало дополнительной лемме и исходной лемме.Вместе с развитием цветочной меристемы, которая достигла большего, чем обычно, размера на более ранней стадии (рис. 2M), эти результаты означают, что эти колоски имели тенденцию давать два цветочка, и детерминированность меристемы колосков была нарушена у mfs1-1. мутант. Сходным образом у мутанта tangari-boushi1 ( tob1 ) у некоторых колосков был дополнительный лемма / палеа-подобный орган между стерильной леммой и исходной леммой (Tanaka et al., 2012). У мутанта snb у некоторых колосков развились избыточные рудиментарные чешуйки, дополнительные лемматические или палеоподобные структуры или боковые соцветия до появления терминальных цветков (Lee et al., 2007). snb + osids1 двойной мутант даже продуцировал больше прицветников-подобных органов, включая рудиментарные чешуйки, чешуйки или палеевидные чешуи, чем отдельные мутанты (Lee and An, 2012). Эти результаты предполагают, что SNB , OsIDS1 , TOB1 и MFS1 регулируют судьбу меристемы колосков, обеспечивая правильное время перехода меристемы колосков в конечную меристему цветков. Напротив, потеря детерминированности меристемы колосков произошла до образования рудиментарной чешуи у мутантов snb и osids1 , но после появления стерильных чешуек у мутантов tob1 и mfs1-1 .Это предполагает, что MFS1 и TOB1 функционируют позже, чем SNB и OsIDS1 . Кроме того, снижение экспрессии SNB и OsIDS1 было обнаружено в мутантных молодых метелках mfs1-1 , что свидетельствует о том, что MFS1 положительно регулирует экспрессию SNB и OsIDS1 .
MFS1 Задает идентичность стерильной леммы
В мутанте mfs1-1 вырожденная стерильная лемма выявила идентичность как стерильной леммы, так и рудиментарной чешуи.У мутанта snb не было обнаружено никаких стерильных лемм на участках, где присутствовали дополнительные рудиментарные чешуйки (Lee et al., 2007), что указывает на гомеотическую трансформацию стерильной чешуи в рудиментарную чешуйку. У мутанта osids1 было показано, что одна из стерильных лемм иногда заменяется рудиментарной чешуей (Lee and An, 2012). SNB и OsIDS1 , как было ранее показано, кодируют белок домена AP2 / ERF (Lee et al., 2007; Lee and An, 2012).Эти результаты предполагают, что MFS1 , SNB и OsIDS1 наделяют важными функциями в развитии стерильной леммы. Сообщалось, что G1 / ELE , OsMADS34 и EG1 определили идентичность стерильной леммы. У этих мутантов стерильная лемма гомеотически трансформировалась в лемму (Li et al., 2009; Yoshida et al., 2009; Gao et al., 2010; Hong et al., 2010; Kobayashi et al., 2010). Эти результаты предполагают, что G1 / ELE , OsMADS34 и EG1 предотвращают преобразование стерильной леммы в лемму, тогда как MFS1 , SNB и OsIDS1 предотвращают стерильную дегенерацию лемма о рудиментарной чешуе.
Существуют две преобладающие гипотезы о происхождении и эволюции бесплодной леммы (Takeoka et al., 1993). Один из них утверждает, что предполагаемый предок рода Oryza имел колоск, который содержал конечный цветочек и два боковых цветочка, которые впоследствии дегенерировали в ходе эволюции, оставив только лемму (Arber, 1934; Kellogg, 2009). по-видимому, произошли от морфологической модификации остатков этой леммы (Yoshida et al., 2009; Kobayashi et al., 2010). Другая гипотеза предполагает, что колосок Oryza spp. имеет только один цветочек, а стерильная лемма и рудиментарная чешуйка повсеместно считаются сильно редуцированными прицветниками (Schmidt, Ambrose, 1998; Terrell et al., 2001; Hong et al., 2010). В мутантах g1 / ele1 , osmads34 и eg1 стерильные леммы были расширены и преобразованы в леммы, что подтверждает первую гипотезу. У мутанта mfs1-1 стерильная лемма выродилась и приобрела идентичность рудиментарной чешуйки, что подтверждает вторую гипотезу.Фактически, у большинства видов трав в колоске отсутствуют стерильные органы, похожие на чешуйки, и есть только один или несколько цветков и прицветниковых чешуек, которые считаются эквивалентами рудиментарных чешуек Oryza spp. (Takeoka et al., 1993; Yoshida et al., 2009; Hong et al., 2010). Прицветник-подобный орган чешуи напоминает лемму по размеру и структуре у некоторых видов трав, таких как кукуруза и пшеница (Kellogg, 2001; Yoshida et al., 2009), тогда как у Oryza spp.(Bommert et al., 2005; Li et al., 2009). Следовательно, лемма, стерильная лемма и рудиментарная чешуйка могут быть гомологичными структурами.
MFS1 Регулирует развитие палеи
Считалось, что у травяных цветов палеа имеет иную идентичность и происхождение от леммы. В целом, палеа считается гомологичной профиллу (первый лист, образованный пазушной меристемой), который формируется на оси цветочка, тогда как лемма соответствует прицветнику (лист, прилегающий к пазушной меристеме), который формируется на колоске. ось (Kellogg, 2001; Ohmori et al., 2009). Недавно появились некоторые свидетельства того, что рисовая палеа — это орган, образованный в результате врожденного слияния боп и мрп, которые потенциально имеют различное происхождение (Francis, 1920; Cusick, 1966; Verbeke, 1992, Zanis, 2007). В частности, во-первых, было показано, что клеточная структура bop очень похожа на структуру леммы, но отличается от структуры mrp (Prasad et al .., 2005; Sang et al .., 2012). Во-вторых, растения с двойной РНКи superwoman1 ( spw1 / osmads16 ) и MADS2 + MADS4 мутантных растений класса B с двойной РНКи показали трансформацию lodicules в органы, похожие на mrp, но не на bop.Более того, мутанты генов арабидопсиса B-класса претерпевают гомеотическую трансформацию лепестков (эквивалентных lodicules) в чашелистики (Nagasawa et al., 2003; Yadav et al., 2007; Yao et al., 2008). Эти находки указывают на то, что mrp, но не bop, гомологичен чашелистнику. У депрессивного мутанта palea1 ( dp1 ), palea, как было показано, замещалось двумя mrp-подобными структурами, и боп был потерян (Luo et al., 2005; Jin et al., 2011). В мутанте с задержкой palea1 ( rep1 ) развитие боп задерживалось, тогда как сверхэкспрессия REP1 вызывала гипердифференцировку клеток mrp (Yuan et al., 2009). У мутанта mfs1-1 боп дегенерировался у большинства цветков и даже отсутствовал в некоторых случаях. Кроме того, недавние исследования показали, что CHIMERIC FLORAL ORGANS1 ( CFO1 / OsMADS32 ) и MOSAIC FLORAL ORGANS1 ( MFO1 / OsMADS6 ) были экспрессированы в мутации mrp, что привело к мутации mrp. mrp в леммоподобные или bop-структуры (Ohmori et al., 2009; Li et al., 2010; Sang et al., 2012). Эти результаты показали, что две части палеи контролируются разными регуляторными путями. В то время как MFS1 , DP1 и REP1 определяют идентичность боп, MFO1 и CFO1 участвуют в регуляции идентичности mrp. В соответствии с этими гипотезами фенотипы палеа mfs1 предполагают, что палеа риса является органом, продуцируемым слиянием mrp и bop, которые имеют разное происхождение.
ЗАКЛЮЧЕНИЕ
В этом исследовании мы охарактеризовали ген риса MFS1 , который принадлежит к кладе с неизвестной функцией в семействе генов AP2 / ERF .У mfs1 колосков были обнаружены лишние леммы, вырожденные стерильные леммы и палеэ. Эти результаты подтверждают, что MFS1 играет важную роль в регуляции детерминированности колосков и идентичности органов. Наши данные также показывают, что MFS1 положительно регулирует экспрессию G1 и IDS1 -подобных генов SNB и OsIDS1 .
МАТЕРИАЛЫ И МЕТОДЫ
Растительные материалы
Два мутанта риса ( Oryza sativa ), mfs1-1 и mfs1-2 , были идентифицированы из обработанного этилметансульфонатом сорта Jinhui 10.cv Jinhui 10 использовали в качестве штамма дикого типа для фенотипического наблюдения. Все растения выращивали на рисовых полях в Чунцине и Хайнане, Китай.
Клонирование на основе карты
MFS1
Мутант mfs1-1 был скрещен с cv Xinong1A, и 1418 растений F2 с мутационным фенотипом были отобраны и использованы в качестве популяции для картирования. Первоначальное картирование генов проводилось с использованием простых маркеров повторов последовательности из общедоступных баз данных по рису, включая Gramene (http: // www.gramene.org) и Программу геномных исследований риса (http://rgp.dna.affrc.go.jp/publicdata/caps/index.html). Затем было выполнено точное картирование с использованием маркеров вставки / делеции, разработанных на основе сравнений геномных последовательностей cv Xinong1A и Jinhui 10 в нашей лаборатории. Последовательности праймеров, используемых при картировании и анализе генов-кандидатов, перечислены в дополнительной таблице S1.
Микроскопический анализ
Метелки собирали на разных стадиях развития и фиксировали в 50% этаноле, 0.9 м ледяной уксусной кислоты и 3,7% формальдегида в течение ночи при 4 ° C, дегидратировали серией градуированных этанолов, пропитывали ксилолом и заливали парафином (Sigma). Срезы толщиной 8 мкм переносили на предметные стекла, покрытые поли-L-Lys, депарафинизировали в ксилоле и дегидратировали с помощью серии этанола. Срезы последовательно окрашивали 1% сафранином (Amresco) и 1% Fast Green (Amresco), затем дегидратировали серией этанола, пропитывали ксилолом и, наконец, помещали под покровное стекло.Световая микроскопия выполнялась на микроскопе Nikon E600. Для сканирующей электронной микроскопии свежие образцы исследовали с помощью растрового электронного микроскопа Hitachi S-3400 с охлаждающим столиком –20 ° C. Стадии раннего развития колосков были такими же, как и определенные ранее (Ikeda et al., 2004).
Выделение РНК
и анализ кПЦР
РНК из корня, стебля, листа, соцветия и молодого цветка выделяли с использованием набора RNeasy Plant Mini от Watson. Первую цепь комплементарной ДНК синтезировали из 2 мкг тотальной РНК с использованием праймеров oligo (dT) 18 в реакционном объеме 25 мкл с использованием набора для обратной транскриптазы SuperScript III (Invitrogen).РНК с обратной транскрипцией (0,5 мкл) использовали в качестве матрицы ПЦР с ген-специфическими праймерами (дополнительная таблица S3). Анализ qPCR выполняли с использованием системы обнаружения последовательностей ABI Prism 7000 и набора SYBR Supermix Kit (Bio-Rad). Было выполнено не менее трех повторов, и были использованы средние значения экспрессии каждого гена.
Гибридизация in situ
Зонд MFS1 длиной 482 п.н. амплифицировали с праймерами MFS1 -F и MFS1 -R и пометили с помощью набора для маркировки DIG RNA Labeling Kit (SP6 / T7) от Roche.Зонды для известных генов цветочных органов были приготовлены с использованием того же метода. Предварительную обработку срезов, гибридизацию и иммунологическое обнаружение проводили, как описано ранее (Sang et al., 2012). Последовательности праймеров перечислены в дополнительной таблице S1.
Конструкция вектора
Для теста комплементации с использованием праймеров MFS1 com- F и MFS1 com-R.Полученные продукты ПЦР расщепляли с использованием Xba I и Eco RI, а затем вставляли в бинарный вектор pCAMBIA1301. Рекомбинантные плазмиды были введены в mfs1-1 с помощью метода трансформации, опосредованной Agrobacterium tumefaciens-, как описано ранее (Sang et al., 2012). Чтобы создать конструкцию для РНКи, мы амплифицировали фрагмент длиной 267 пар оснований комплементарной ДНК MFS1 с праймерами MFS1, Ri-F ( Spe I, Kpn I) и MFS1 Ri-R ( Sac I, Bam HI), как показано в дополнительной таблице S1.Полученные продукты ПЦР сначала расщепляли с использованием Spe I и Sac I, а затем лигировали в вектор pTCK303 (Wang et al., 2004) для получения промежуточного вектора. Затем продукты ПЦР расщепляли с использованием Kpn I и Bam HI и лигировали в промежуточный вектор. Рекомбинантные плазмиды трансформировали в растения Zh21 с помощью метода трансформации, опосредованной A. tumefaciens-. Последовательности праймеров перечислены в дополнительной таблице S1.
Субклеточная локализация
Кодирующая область MFS1 без стоп-кодона была амплифицирована с использованием пары праймеров MFS1 OE-F и MFS1 OE-R, которые содержат сайты Xba, I и Bam HI, соответственно. (Дополнительная таблица S1).Фрагмент был клонирован в кассету экспрессии 35S-GFP (S65T) -NOS (pCAMBIA1301) с соответствующими модификациями, в результате чего был получен вектор слияния MFS1 -GFP. Плазмиды GFP и MFS1 -GFP трансформировали в протопласты риса, как описано ранее (Li et al., 2009). Через 8–16 ч инкубации при 28 ° C флуоресценцию GFP наблюдали с помощью микроскопа Nikon E600.
Последовательности белков и филогенетический анализ
Последовательности белков были получены путем поиска в GenBank (http: // www.ncbi.nlm.nih.gov/genbank/), используя последовательность MFS1 в качестве запроса. Филогенетическое дерево было построено с использованием MEGA 5.0 (Tamura et al., 2011). Дерево было построено с использованием метода максимального правдоподобия на основе модели на основе матрицы JTT с самыми низкими оценками байесовского информационного критерия (Jones et al., 1992; Tamura et al., 2011). Значения поддержки начальной загрузки для каждого узла из 500 реплик показаны рядом с ветвями (Felsenstein, 1985). Исходное дерево для эвристического поиска было получено автоматически следующим образом.Когда количество общих сайтов было меньше 100 или меньше четверти от общего количества сайтов, использовался метод максимальной экономии; в противном случае использовался метод соединения биососедов с матрицей кластерных расстояний Маркова. Дискретное γ-распределение использовалось для моделирования различий в скорости эволюции между участками (пять категорий [+ G ], параметр = 0,6362). Дерево было нарисовано в масштабе, с длиной ветвей, измеренной количеством замен на сайт.
Данные последовательности из этой статьи можно найти в библиотеках данных GenBank / EMBL под номерами доступа: SNB (ABD24033), OsIDS1 (NM_001058244), G1 / ELE (AB512480), DL (AB106553) (AB106553) , OsMADS1 (NM_001055911), OsMADS6 (FJ666318), OsMADS14 (NM_001057835) и OsMADS15 (NM_001065255).
Дополнительные данные
Следующие материалы доступны в онлайн-версии этой статьи.
Сноски
Автор, ответственный за распространение материалов, составляющих выводы, представленные в этой статье, в соответствии с политикой, описанной в Инструкциях для авторов (www.plantphysiol.org): Guanghua He (hegh2968 {at} yahoo .com.cn).
www.plantphysiol.org/cgi/doi/10.1104/pp.113.216044
↵1 Работа была поддержана Национальным фондом естественных наук Китая (грант №31071071), Китайская программа 863 (грант № 2011AA10A100), проект «111» (грант № B12006), Крупные исследовательские проекты Чунцина, Китай (грант № 2012ggc80002), а также Фонды фундаментальных исследований для Центральной Университеты, Китай (грант № XDJK2010C073).
№2 Эти авторы внесли одинаковый вклад в статью.
↵ [C] Некоторые рисунки в этой статье отображаются в Интернете в цвете, а в печатном издании — в черно-белом.
↵ [W] Онлайн-версия этой статьи содержит данные только для Интернета.
Глоссарий
- bop
- тело палеи
- mrp
- маргинальные области палеи
- кПЦР
- количественная обратная транскрипция-ПЦР
- ERNAi
- 4
- 4
- 4
- 492 RNAi
- 4 924 / EREBP
- 4
24
- Получено 17 января 2013 г.
- Принято 26 апреля 2013 г.
- Опубликовано 29 апреля 2013 г.
ССЫЛКИ
- ↵
- ↵
Arber A (1934) Gramineae: Исследование злаков, бамбука и трав.Cambridge University Press, Кембридж, Великобритания
- ↵
- ↵
- ↵
- ↵
Clifford HT (1987) Морфология колосков и цветков. В Т. Р. Содерстрем, К. В. Хилу, К. С. Кэмпбелл, М. Е. Баркворт, ред., «Систематика и эволюция травы». Smithsonian Institution Press, Вашингтон, округ Колумбия, стр. 21–30
- ↵
- ↵
- ↵
- ↵
- ↵
Cusick F (1966) О филогенетических и онтогенетических слияниях. In EG Cutter, ed, Trends in Plant Morphogenesis. Longmans, Green, London, pp 170–183
- ↵
- ↵
- ↵
Francis ME (1920) Book of Grasses. Doubleday, Page, Garden City, NY
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
Malcomber ST, Preston JC, Reinheimer R, Kossuth J, Kellogg EA (2006) Эволюция генов развития и происхождение разнообразия соцветий травы In J Leebens-Mack, Д. Е. Солтис, П. С. Солтис, редакторы, «Генетика развития цветка».Academic Press, New York, pp. 383–421
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
Takeoka Y, Shimizu M, Wada T (1993) Panicles. In T Matsuo, K Hoshikawa, ред., Science of the Rice Plant, Vol I. Nobunkyo, Tokyo, pp 295–326
- ↵
- ↵
- ↵
- ↵
- ↵
Weberling F. (1989) Морфология цветов и соцветий.Cambridge University Press, Кембридж, Великобритания
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
Неопределенный ген апекс1 цветков регулирует детерминированность и идентичность меристемы в соцветиях кукурузы
Резюме
Меристемы могут быть как определенными, так и неопределенными. У кукурузы из недетерминантной меристемы соцветия образуются три типа детерминированных меристем: пара колосков, колоски и цветочные меристемы.Эти меристемы определяются своим положением и продуктами. Мы обнаружили ген у кукурузы, неопределенной вершины цветка ( ifa1 ), который регулирует детерминированность меристемы. Дефект, обнаруженный у мутантов ifa1 , специфичен для меристем и не влияет на боковые органы. У мутантов ifa1 детерминированные меристемы становятся менее детерминированными. Меристема пары колосков инициирует больше, чем пара колосков, а меристема колосков инициирует больше, чем два нормальных цветка.Цветочная меристема правильно инициирует все органы, но зачаток семяпочки, конечный продукт цветочной меристемы, увеличивается и разрастается, экспрессируя как гены-маркеры меристемы, так и семяпочки. Роль ifa1 в идентичности меристемы в дополнение к детерминированности меристемы была выявлена с помощью анализа двойных мутантов. В двойных мутантах zea agamous1 ( zag1 ) ifa1 женская цветочная меристема превращается в меристему ветвления, тогда как мужская цветочная меристема превращается в меристему колосков.У недетерминантных spikelet1 (ids1) ifa1 двойных мутантов женские меристемы колосков превращаются в меристемы ветвей, а мужские меристемы колосков превращаются в меристемы пары колосков. Фенотипы с двойными мутантами предполагают, что спецификация меристем в соцветии кукурузы включает отдельные шаги в интегрированном процессе.
ВВЕДЕНИЕ
Развитие растений во многом зависит от активности меристем, пулов недифференцированных клеток, которые делятся для поддержания меристемы и обеспечения клеток для боковых органов, ветвей и стеблей (Steeves and Sussex, 1989).Неопределенные меристемы непрерывно производят органы до старения, тогда как детерминированные меристемы перестают производить новые клетки после того, как инициируется фиксированное количество зачатков. Меристемы соцветий часто служат примером неопределенных меристем, производя неопределенное количество цветочных меристем. Цветочные меристемы обычно детерминантные; после инициирования четырех оборотов цветковых органов они расходуются на производство плодолистиков. Кукуруза и другие родственные травы представляют собой отличную систему для изучения перехода от неопределенной меристемы к определенной благодаря наличию промежуточных стадий ветвления до образования цветков.
Расположение цветочных меристем на соцветиях широко варьирует. У некоторых видов цветочные меристемы рождаются непосредственно на соцветии, а у других возникают из боковых ветвей. Расположение и филлотактический рисунок цветочных меристем также могут обеспечить большое разнообразие форм. Многие виды переходят от одного филлотактического паттерна на вегетативной стадии к другому паттерну цветения. Меристема соцветий кукурузы дает зачатки в виде многоядерных (или многорядных) структур.Ранние зародыши развиваются в меристемы ветвей, а сформировавшиеся позже зачатки становятся меристемами пар колосков (рис. 1). Меристема ветвей (ВМ) похожа на меристему соцветия в непрерывном образовании меристем пар колосков (SPM), но отличается схемой зарождения, которая является двурядной или двурядной. ВП образуют две меристемы колосков (СМ), каждая из которых образует две цветочные меристемы (рис. 1А). Как однодомное растение кукуруза имеет два типа меристемы соцветий: верхнее мужское соцветие (кисточка, рис.1D) и бокового женского соцветия (колос, рис. 1E) (Cheng et al., 1983; McSteen et al., 2000). Кисточка образуется в результате превращения вегетативной меристемы в соцветие, а колос возникает из пазушной меристемы вегетативного листа. И кисточка, и ухо производят SPM, но производство BM в ухе подавляется.
Рис. 1.
Развитие соцветий кукурузы. (A) Различные типы меристемы кукурузы. Меристема соцветия (IM) образует меристемы пары колосков (SPM), которые образуют две меристемы колосков (SM), каждая из которых образует две меристемы цветков (FM).Кисточки также образуют в основании ветвистые меристемы (БМ). (B, C) Схема пары колосков. На каждом колоске две чешуи и два цветочка. Зрелый цветочек состоит из чешуи, палеи, трех лодикул (одна из которых рудиментарная и не показана), трех тычинок и пестика. Пестик прерывается при мужских колосках (B). У самок колосков прерываются все тычинки и нижний цветочек (С). (D) Мужское соцветие. Базальные ответвления на кисточке образуют неопределенные BM, которые по типу образуют SPM.SPM порождают два SM, которые находятся бок о бок. Каждая SM образует две цветочные меристемы (FM). (E) Развитие женского соцветия происходит аналогично развитию мужского соцветия, только BM подавляются в ухе. Область в рамке содержит шесть колосков. В каждом колоске видны верхняя ЖМ и внешняя чешуйка. Нижняя ФМ и внутренняя чешуя не видны.
Как и меристема цветков, SPM и SM детерминированы, давая фиксированное количество производных. Ряд мутаций кукурузы влияет на детерминированность этих меристем.У мутантов ramosa 1 (ra1) SPM на главной оси кисточки становятся неопределенными, давая множественные колоски (Postlethwait and Nelson, 1964; Veit et al., 1993). Сами SM подвержены ряду мутаций, включая неопределенных колосков1 ( ids1 ), Tasselseed6 ( Ts6 ) и обратной ориентации зародышей1 ( rgo1 ) (Irish, 1997; Chuck et al., 1998; McSteen et al., 2000). В ids1 , Ts6 и rgo1 появляются дополнительные цветки, что указывает на неопределенность SM.Детерминация самой цветочной меристемы затрагивается у кукурузы AGAMOUS гомолог, zea agamous1 ( zag1 ), узловатый1 ( kn1 ) и толстый кистевидный карлик 1 ( td1a) ., 1996; Kerstetter et al., 1997; McSteen et al., 2000). У каждого из этих трех мутантов появляется больше плодолистиков.
Несколько мутаций кукурузы также влияют на идентичность продуктов меристемы. У мутантов tasselseed4 (ts4) SPM инициируют SPM и, таким образом, идентичны BM (Irish, 1997).В разветвленных silkless1 ( bd1) мутантных ушах идентичность SM изменена и цветочные меристемы не продуцируются (Kempton, 1934; Colombo et al., 1998). В indeterminate1 (id1) SM превращается в вегетативную меристему после образования двух цветков (Galinat and Naylor, 1951) (D. L.C. и J. Colasanti, неопубликовано).
Описанные ранее мутации затрагивают только один конкретный тип меристемы. Здесь мы описываем новую мутацию undeterminate flower apex1 ( ifa1 ), которая затрагивает все детерминированные меристемы соцветия кукурузы; количество колосков и цветков увеличивается, зачатки семяпочек разрастаются.Двойные мутанты с zag1 и ids1 обнаруживают дополнительную роль ifa1 в идентичности меристемы. Спецификация идентичности меристемы различна у мужских и женских цветков.
МАТЕРИАЛЫ И МЕТОДЫ
Генетика
Пыльцу растений, гомозиготных по ifa1-r (r для справки), скрещивали со стандартным набором из восковых линий реципрокной транслокации с меткой (Laughnan and Gabay-Laughnan, 1993).Нарушение сегрегации с использованием T1-9c продемонстрировало свидетельство сцепления, предполагая, что ifa1-r картирован на хромосоме 1. Несколько маркеров RFLP были использованы для дальнейшего определения местоположения ifa1 на хромосоме 1S. ifa1-r картировал 3 см от umc76 и менее 1 см от zmm14 (ноль рекомбинантов в 110 хромосомах). ifa1-r был интрогрессирован от четырех до шести поколений в несколько инбредных линий, включая B73, Mo17, W23, A188, A619 и A632.
zag1-mum1 был подарком Р. Шмидта (Калифорнийский университет, Сан-Диего). zag1, и ifa1. Двойные мутанты были получены путем скрещивания пыльцы растений, гомозиготных по ifa1-r , с колосьями zag1-mum1 / + . Растения F 1 были самоопыляемыми, а потомство F 2 оценивали на сегрегацию обоих фенотипов ifa1-r и zag1 в семействе. Генотип двойных мутантов подтверждали скрещиванием предполагаемых двойных мутантных растений как мужских с растениями, гетерозиготными по zag1 и ifa1-r , или путем оценки диагностических маркеров RFLP, связанных с каждой мутацией.Двойные мутанты ifa1-r с id1-mum1 (Chuck et al., 1998) были сконструированы аналогичным образом. Аллели, используемые при конструировании двойных мутантов, вероятно, будут нулевыми, исходя из того факта, что растений ifa1-r / — аналогичны растениям ifa1-r / ifa1-r при сравнении на том же фоне, и что ген-специфичный транскрипты отсутствуют у ids1-mum1 и zag1-mum1 (Mena et al., 1996; Chuck et al., 1998).
Микроскопия
Молодые зачатки уха, используемые для экспериментов с РНК in situ и для сканирующей электронной микроскопии, были собраны с растений, выращенных в теплице.Гибридизацию in situ проводили, как описано (Jackson et al., 1994), с использованием кДНК kn1 или zag2 в качестве зондов. Фрагмент длиной 650 п.н. ниже бокс-домена MADS был амплифицирован из субклона кДНК zag2 с использованием следующих пар праймеров: 5′-TAA TAC GAC TCA CTA TAG GCC TTA ACC TGA TCA CAG CCT-3 ‘и 5’-ATT TAA CCC TCA CTA AAG GCA TCT CTA AGA TCA GGG-3 ‘, где подчеркнутые последовательности представляют собой промоторные последовательности Т7 и Т3 соответственно. Зонды были созданы путем транскрипции in vitro с использованием полимеразы Т7 для создания антисмысловой цепи и полимеразы Т3 для создания смысловой цепи.Молодые зачатки ушей фиксировали в FAA (50% этанол, 5% уксусная кислота, 3,7% параформальдегид) при 4 ° C в течение ночи и дегидратировали в серии этанола с постепенным изменением концентрации до 100%. Образцы были высушены до критической точки и напылены палладием. Образцы просматривали на растровом электронном микроскопе ISI 30 при ускоряющем напряжении 10 кВ.
РЕЗУЛЬТАТЫ
ifa1 отображается на хромосому 1
ifa1-r (r для справки) был идентифицирован в линии кукурузы, несущей транспозон Ac , но не связан с Ac (данные не показаны).Мы сопоставили его с хромосомой 1 с помощью восковых линий транслокации (Laughnan and Gabay-Laughnan, 1993) и показали, что он ведет себя как одиночная рецессивная мутация. Скрещивание ifa1 — r с линиями транслокации BA, которые приводят к хромосомным дефектам (Beckett, 1993), подтверждают его положение на коротком плече хромосомы 1. Дальнейшее картирование с использованием маркеров RFLP размещает ifa1 в пределах 3 см от umc76 и менее 1 см от zmm14 .
Фенотип растений, несущих аллель ifa1-r , обнаруженный дефицитом, аналогичен гомозиготам ifa1-r , что позволяет предположить, что ifa1-r является нулевой мутацией. ifa1-r был скрещен с несколькими инбредными линиями и является полностью пенетрантным. Аспекты фенотипа более выражены в исходных линиях, в которых он был идентифицирован, чем после интрогрессии в данный инбред. Дополнительные аллели были обнаружены путем скрининга популяций F -1 на наличие аллелей, индуцированных транспозоном и EMS. Двойные гетерозиготы не так серьезны, как гомозиготы ifa1-r . В этой статье сообщается о фенотипе ifa1-r .
ifa1 регулирует детерминированность пары колосков, меристем колосков и цветков
Меристемы соцветий кисточки и колоса образуют меристемы пары колосков (SPM) в виде полистихового разветвления (рис.1D, E). SPM продуцируют пару меристем колосков (SM). В кисточке верхний колоск, имеющий более длинную ножку, называется колоском на ножке, а нижний колоск сидячий (рис. 2А). Каждый SM дает четыре листообразных органа по определенному образцу. Первые две, также известные как чешуйки, бесплодны. Следующие два (называемые леммой) плодовиты и содержат цветочные меристемы в пазухах (рис. 1B, C). Лемма и продукты цветочной меристемы составляют цветочек. Таким образом, колоск состоит из пары чешуек, охватывающих два цветочка (рис.1B, рис. 2B).
Рис. 2.
мутанты ifa1 дают дополнительные колоски и дополнительные соцветия. (A) Цветоножка и сидячий колоск от обычной кисточки. (B) Мужской колоск с двумя цветочками в паре чешуек. (C) У мутантов ifa1 более двух цветков наблюдается в колоске. (D) Вместо пары колосков образуются три или более колосков. г, чешуйка; ф, цветочек; зр, колоск; sp *, дополнительный колоск, p, цветоножка.
У растений ifa1-r SPM и SM становятся менее детерминированными, производя больше колосков и цветков (рис. 2). Дополнительные колоски, как и обычные колоски, содержат соцветия, окруженные чешуей. Дополнительные колоски в кисточке имеют ножки и начинаются последовательно. Гомозиготные мутанты ifa1-r могут давать более двух цветков на один колосок, которые, как и нормальные соцветия, разделены короткими междоузлиями и содержатся внутри двух стерильных чешуек (рис.2С). Лишние колоски и соцветия встречаются не так часто после многократных обратных скрещиваний в один инбред, и их больше можно найти на кончике основной кисточки и кончиках боковых ветвей. Образование дополнительных колосков и цветочков у мутантов ifa1 предполагает, что ifa1 регулирует детерминированность SPM и SM, соответственно.
Начальные этапы развития мужских и женских цветков схожи, на более поздних стадиях они дифференцируются в однополые цветки.Первым органом, образованным из цветочной меристемы, является палеа, перепончатый листообразный орган, за которым следуют три лодикулы (одна подавленная), три тычинки и пестик (рис. 1B, C). Лодикулы считаются рудиментарными лепестками на основании экспрессии маркерных генов (Ambrose et al., 2000). Пестик состоит из трех плодолистиков (одного рудиментарного и двух сросшихся), которые удлиняются, образуя шелк (рис. 3А) (Никерсон, 1954). Внутри сросшихся плодолистиков находится единственная семяпочка, состоящая из покровов и нуцеллярной ткани, окружающей женский гаметофит (рис.3Б). Пестик прерывается на ранней стадии развития цветков кисточки (Рис. 1B). В ухе все тычинки, а также нижний цветочек каждого колоска прерываются, в результате чего на каждом колоске образуется один женский цветок (Рис. 1C).
Рис. 3.
Цветочный фенотип мутантов ifa1 . (A) Нормальный пестик. Два плодолистика сливаются и удлиняются, образуя шелк. (B) Зачаток семяпочки обнажается внутри сросшихся плодолистиков. (C) ifa1-r пестик.Нуклеароподобная пролиферативная ткань возникает от верхушки цветка, видны остатки стенки плодолистика и рудиментарный шелк. (D) Иногда в центре пестика ifa1-r образуются множественные семяподобные выступы. (E) ifa1-r колоск с эктопическим соцветием от обоих соцветий. Этот фенотип встречается с низкой частотой. (F) Нормальный мужской колоск. Видны тычинки верхнего цветочка, тычинки нижнего цветочка остаются в окружении чешуи и палеи.(G) ifa1-r мужской цветочек. Внутри оборотной части тычинки присутствует нуклеароподобная пролиферативная ткань (стрелка). (H) ifa1-r мужской цветочек с дополнительными тычинками в центре тычинки. (I) Рассеченный цветочек из H. Внутри тычинки находятся листообразные органы и несколько тычинок. си, шелк; op, зачаток яйцеклетки; с, тычинка; lf, нижний цветочек.
У мутантов ifa1 цветочная меристема становится менее детерминированной. Женские цветки характеризуются короткими шелками и неразвитыми плодолистиками, которые окружают массу выступающей нуцеллусоподобной ткани (рис.3С). Идентичность ткани как продукта цветочной меристемы определялась по ее расположению внутри оборотов абортированных тычинок (рис. 3D). Фенотип торчащего нуцеллуса наиболее выражен на кончике уха. Иногда можно увидеть более одной выступающей нуцеллярной массы (рис. 3D) или зачаток семяпочки, по-видимому, трансформируется в структуру соцветия (рис. 3E). Эти преобразования наблюдались в исходных линиях транспозона Ac и происходят при значениях менее 0.1% времени в любом интрогрессивном материале. Фенотип выступающего нуцеллуса является 100% пенетрантным, когда ifa1-r интрогрессирован во все протестированные инбредные породы.
Аналогичный фенотип наблюдался у цветков мужских колосков. У нормальных цветков три тычинки при созревании обнаруживаются внутри вмещающей палеи и леммы (рис. 3F). У мутантов ifa1 увеличенная нуцеллярная масса иногда образуется внутри оборотов трех тычинок (Fig. 3G). У цветков, расположенных ближе к кончику кисточки или веточки кисточки, в центре цветка вместо увеличенной нуцеллярной массы образуются дополнительные тычинки (рис.3H). Каждая из дополнительных тычинок, по-видимому, развивается в пазухе леммоподобной структуры (Рис. 3I), что указывает на то, что дополнительные тычинки возникают не из-за образования дополнительных оборотов тычинок, а скорее из-за устойчивой цветочной меристемы. Нуцеллярная масса или лишние тычинки обнаруживаются как у верхних, так и у нижних мужских цветков. Таким образом, как мужские, так и женские цветочные меристемы продолжают размножаться. Этот результат предполагает, что ifa1 регулирует детерминацию цветочной меристемы.
Сканирующая электронная микроскопия выявляет ранние события у мутантов
ifa1
Чтобы определить, когда цветков ifa1-r впервые отклоняются от нормы, мы исследовали развивающиеся женские соцветия от мутантов (рис.4F-I) и их нормальных братьев и сестер (рис. 4A-D) с помощью сканирующей электронной микроскопии (SEM). Поскольку у ifa1-r формирование тычинок не затрагивается, мы использовали стадию развития тычинок в качестве стандарта для сравнения развивающихся соцветий. На рис. 4A, F инициируются как верхняя, так и нижняя цветочные меристемы. Хотя нижняя цветочная меристема инициируется первой (Cheng et al., 1983), она отстает в развитии. Нормальные цветочные меристемы и меристемы ifa1-r кажутся похожими как по размеру, так и по морфологии, когда зарождаются зачатки тычинок.Нормальные соцветия образуют гребень из ткани плодолистика, который возникает на адаксиальной стороне (относительно меристемы соцветия) цветочной меристемы. Плодолистики окружают цветочную меристему, которая на этой стадии окончательно дифференцируется в зачаток семяпочки (рис. 4B, C) и удлиняется, образуя шелк (рис. 4D).
Рис. 4.
СЭМ нормальных и ifa1-r колосков и цветочков. (A-D) Нормальный цветочек. (F-I) ifa1-r цветочек. (A, F) Верхние FM инициировали три тычинки, одна из которых скрыта.Часть чешуи удаляется с колоска в F, чтобы обнажить большую часть нижней FM. (B, G) Гребень плодолистика формируется на адаксиальной стороне FM. (C, H) Гребень плодолистика начинает охватывать зачаток семяпочки у нормальных (C), но не у мутантов ifa1 (H). (D) плодолистики почти полностью охватили семяпочку. (I) Зачаток семяпочки у мутантов ifa1 формирует органы по двоякой схеме. Нижняя ЧМ тоже ненормальная. (E) Колоски у нормальных растений развиваются парами, при этом внешние чешуйки образуются на абаксиальной стороне.Верхний цветочек, развивающийся впереди нижнего цветочка, формируется на адаксиальной стороне колоска. И внутренняя чешуйка, и нижний цветочек не видны. (F) У мутантов ifa1 между парой колосков образуется дополнительный колоск. Этот дополнительный колоск развивается в обратной ориентации, так что внешняя чешуйка (см. Маленькую стрелку) является адаксиальной, а внутренняя чешуйка и гребень плодолистика верхнего цветочка (см. Большая стрелка) — абаксиальными. lf — нижний цветочек; ул — тычинка; op, зачаток яйцеклетки; ог, наружная чешуя; ig, внутренняя чешуйка.
У мутантов ifa1 гребень плодолистика не охватывает зачаток семяпочки (рис. 4G). Увеличенный и деформированный зачаток семяпочки дает дополнительные органы в виде дистиха (рис. 4H, I). Предполагается, что эти органы представляют собой плодолистики из-за утолщенного гребня. Нижняя цветочная меристема также поражена у мутантов ifa1 . На рис. 4I нижняя цветочная меристема дала один плодолистик и дает второй в противоположном положении.
Мы также исследовали происхождение дополнительных колосков у мутантов ifa1 с помощью SEM. Как и у нормальных растений, SPM делится неравномерно, образуя два зачатка. У ifa1-r более крупный зачаток снова делится на два неравных зачатка, один из которых развивается в дополнительный колоск (данные не показаны). На рис. 4E показана пара колосков от нормального человека. Верхние цветочные меристемы уже положили начало тычинкам и пестикам. У мутанта ifa1 между парой колосков появляется дополнительный колоск (рис.4J). Между двумя нормально расположенными колосками всегда находились лишние колоски (данные не показаны). Этот дополнительный колоск имеет противоположную ориентацию по отношению к другим колоскам, что видно по ориентации гребня плодолистика на верхнем цветке (стрелка на рис. 4J). Гребень плодолистика, который обычно является адаксиальным по положению, теперь абаксиальный. Дополнительный колоск, по-видимому, немного задерживается в развитии по сравнению с другими колосками. Этот результат свидетельствует о том, что дополнительный колоск возникает скорее из-за ветвления, чем из-за подразделения.
Таким образом, SEM показывает, что многие аспекты нормального развития сохраняются у мутанта ifa1 . SPM создает SM, которые создают FM, которые инициируют правильный набор органов. Однако в каждом случае определенные меристемы производят больше продуктов, чем обычно. Разные образцы инициации предполагают, что дефект связан с определенностью, а не размером меристемы.
Экспрессия маркерных генов плодолистика и меристемы у
ifa1 мутантов
Чтобы оценить детерминированность меристем ifa1 и на молекулярном уровне, мы провели гибридизацию in situ с геном гомеобокса, узловых1 ( kn1 ) (Smith et al., 1992; Джексон и др., 1994). kn1 экспрессируется во всех меристемах и осях побегов и снижается по мере образования боковых органов. Он выражен в паре колосков, меристемах колосков и цветков, но не в чешуях или других цветочных зачатках. Как показано на фиг. 5A, экспрессия kn1 в этих колосках отступает от верхней меристемы цветков по мере развития зачатка семяпочки. В то же время экспрессия kn1 все еще сильна в нижней меристеме цветка, которая находится в абаксиальном положении.Позднее в развитии ни шелк, ни семяпочка не экспрессируют kn1, , хотя основание вершины цветка и недифференцированная нижняя меристема цветка продолжают это делать (Fig. 5D).
Рис. 5.
kn1 Экспрессия гена в колосках и цветочках уха. (A) Экспрессия kn1 в развивающихся колосках. На продольном срезе нормального соцветия виден только один колоск из пары колосков. Нижний цветочек формируется на абаксиальной стороне каждого колоска.(B) Экспрессия kn1 в развивающихся колосках ifa1-r . На продольном срезе видна часть дополнительного колоска (отмечена *), которая перекрывается с развивающейся парой колосков. Нижний цветочек с адаксиальной стороны развивается дополнительными колосками. (C) Экспрессия kn1 в эктопическом костном мозге, возникающая из уха ifa1-r zag1 . (D) kn1 не экспрессируется в шелке или семяпочке нормальных растений. Экспрессия обнаруживается в нижней цветочной меристеме и цветочной оси ниже семяпочки.(E-H) Серийный срез зачатка мутантной яйцеклетки ifa1 . kn1 также подавляется при инициации зачатка семяпочки, однако, как показано на серии срезов по 10 мкм от той же верхушки цветка, kn1 экспрессируется в небольшой группе клеток в центре зачатка. уф, верхний цветочек; lf — нижний цветочек; *, лишний колоск; яйцеклетка; op, зачаток яйцеклетки; ca, плодолистник; ов, яйцеклетка.
Характер экспрессии kn1 в продольном срезе соцветия ifa1-r показывает наличие лишнего колоска (* на рис.5Б). В нормальном соцветии пары колосков развиваются бок о бок, поэтому на продольном срезе можно наблюдать только один столбик колосков (рис. 5А). Наличие дополнительного колоска можно наблюдать на продольном срезе соцветия ifa1-r , потому что часть дополнительного колоска перекрывается с нормальной парой колосков во время развития (рис. 5B). От одного из этих лишних колосков ответвляется молодая цветочная меристема. По стадии своего развития эта расположенная адаксиально меристема является нижней меристемой цветка.Это наблюдение согласуется с нашим SEM соцветия ifa1-r (рис. 4J), что указывает на то, что ориентация дополнительного колоска противоположна ориентации нормальных колосков.
Мы исследовали экспрессию kn1 в зачатках семяпочек ifa1-r цветков. В цветочных меристемах дикого типа kn1 экспрессируется до тех пор, пока не инициируются зачатки семяпочки (фиг. 5D). На стадии, когда гребень плодолистика начинает удлиняться, экспрессия kn1 формирует резкую линию между зачатком семяпочки и осью цветочной меристемы.У мутантов ifa1 экспрессия kn1 также ограничена меристемами побегов и не обнаруживается в органах цветка. Экспрессия исчезает из зачатка яйцеклетки, как у нормальных братьев и сестер, однако экспрессия возвращается в нескольких клетках в центре увеличивающейся структуры, подобной яйцеклетке. Последовательный срез зачатка яйцеклетки ifa1-r обнаруживает небольшую группу центрально расположенных клеток, которые экспрессируют kn1 (фиг. 5E-H). Этот паттерн экспрессии наблюдали в нескольких секционированных семязачатках (данные не показаны).Таким образом, ifa1 действует прямо или косвенно, отрицательно регулируя kn1 в подмножестве клеток в центре зачатка яйцеклетки.
Поскольку экспрессия kn1 исчезает в зарождающихся зачатках семяпочки, экспрессируются гены, специфичные для плодолистиков и семяпочек, такие как zea agamous2 ( zag2 ) (Schmidt et al., 1993). Мы использовали zag2 в качестве зонда для гибридизации in situ, чтобы определить, приобрел ли примордий ifa1-r яйцеклетку идентичность . Как и в женских цветках дикого типа, мРНК zag2 была обнаружена в зачатке семяпочки и внутренней стенке плодолистика пестика ifa1-r (фиг. 6A, B). Хотя зачаток ifa1-r значительно увеличен по сравнению с нормальными яйцеклетками, экспрессия zag2 все еще видна. Мы также наблюдали мегаспоровые материнские клетки в некоторых зачатках яйцеклеток ifa1-r (данные не показаны). Эти результаты показывают, что меристема цветков у мутантов ifa1 приобрела идентичность, подобную яйцеклетке.Таким образом, экспрессия kn1 в некоторых клетках зачатка яйцеклетки ifa1-r происходит не просто из-за неспособности дифференцироваться в ткань яйцеклетки. По-видимому, субнабор ifa1-r клеток зачатка яйцеклетки восстанавливает или сохраняет меристематическую идентичность.
Рис. 6.
Экспрессия гена
zag2 в нормальном цветке и ifa1-r цветка. (A) zag2 экспрессируется в семяпочке и стенке плодолистика. (B) У ifa1 мутантов , zag2 также экспрессируется в стенке плодолистика и инициирующем зачатке семяпочки.ca, плодолистник; op, зачаток яйцеклетки.
Генетическое взаимодействие
ifa1-r с мутантами, влияющими на детерминированность цветков и колосков
Чтобы получить более полное представление о функции ifa1 в детерминированности меристемы, мы сконструировали двойные мутанты между ifa1-r и двумя другими мутациями, которые влияют на детерминированность меристемы цветков и колосков, zag1-mum1 и ids1-mum1 . Мутации в zag1 , гомологе кукурузы AGAMOUS (Yanofsky et al., 1990) в Arabidopsis и PLENA в Antirrhinum (Bradley et al., 1993), приводят к потере детерминированности женской цветочной меристемы (Mena et al., 1996). zag1 мутантные цветки образуют лишние плодолистики и лишние шелки (рис. 7B) и иногда образуют массу ткани в мутовке плодолистика. Мутации в zag1 не влияют на морфологию мужских цветков.
Рис. 7.
ifa1 zag1 фенотип двойного мутанта.(A) ifa1-r ухо. (B) zag1-mum1 ухо. (C) ifa1-r zag1-mum1 колос показывает соцветия, выходящие из центра каждого цветочка. (D) Крупный план (C). (E) Обычная кисточка. (F-H) ifa1-r zag1-mum1 кисточка. (G) ifa1-r zag1-mum1 цветочек показывает эктопический колоск, выходящий из центра тычинки, где должен был быть прерванный пестик. (H) Внематочный колоск G был открыт, чтобы выявить стаминоидные соцветия, возникающие из пазух листоподобных органов.Стрелки в G и H указывают на одну из лодикул (lo).
Женские цветки двойных мутантов ifa1 zag1 обнаруживают фенотип, не обнаруженный ни у одного из одиночных мутантов (рис. 7A, B). Все женские цветки растений с дефектом ifa1 и zag1 развивают ветвящуюся структуру в центре цветка (рис. 7C, D). Ветвящиеся структуры образуют пары колосков по типу двоякой структуры и поэтому считаются BM. Эти вторичные BM возникают в обороте тычинок (данные не показаны).Цветки, продуцируемые вторичным BM, повторяют дефектный фенотип, давая начало третичным ветвям. В соответствии с этим неопределенным фенотипом, kn1 экспрессируется в SPM, которые возникают из этого эктопического BM (Fig. 5C). Фенотип полностью пенетрантный и очень выразительный как на инбредном, так и на смешанном фоне (данные не показаны).
Кисточки двойных мутантов ifa1 zag1 (рис. 7F) четко отличаются от кисточек zag1-mum1 , которые имеют нормальную морфологию, и кисточек ifa1-r , которые немного более густые (рис.7E, F). Отделение цветочка от двойной мутантной кисточки, показанной на рис. 7G, выявляет эктопический колоск в обороте тычинки. Этот колоск находится в положении пестика, который обычно прерывается у мужских цветков. У эктопического колоска листовые чешуи, которых нет у нормальных колосков. Когда чешуйчатые чешуи эктопического колоска открываются, выявляются несколько тычинок, окруженных дополнительными листовидными органами (рис. 7H). Этот фенотип предполагает, что эктопический колоск образовал несколько цветков.Таким образом, ifa1 и zag1 независимо регулируют детерминированность цветочной меристемы, но действуют избыточно, регулируя идентичность цветочной меристемы. Когда оба гена ifa1 и zag1 мутируют, меристема цветков превращается в меристему ответвления в ухе и в меристему колосков в кисточке.
неопределенный spikelet1 (ids1), , который кодирует AP2-подобный белок, контролирует детерминированность меристемы колосков у кукурузы (Chuck et al., 1998). Аллель ids1-mum1 приводит к образованию множества цветков из меристемы колосков. Этот фенотип подобен одному аспекту фенотипа ifa1-r (Fig. 2C). У мутантов ids1 поражаются как женские, так и мужские соцветия. Цветочки в ухе образуют шелк (рис. 8A), но обычно бесплодны (Chuck et al., 1998).
Рис. 8.
ifa1 ids1 фенотип двойного мутанта. Индивидуумы происходят из семьи, разделяющей по фенотипу двойных мутантов ifa1 ids1 .(A) ids1-mum1 ухо. (B) ifa1-r ухо. (C-F) ifa1-r ids1-mum1 ухо. Каждый колоск образует эктопическое соцветие. (E) ifa1-r ids1-mum1 пара колосков. Поражаются оба колоска в паре. (F) ifa1-r ids1-mum1 колоск с тремя цветочками и эктопическим соцветием. Внематочное соцветие образует по своей оси пары колосков (стрелка указывает на пару колосков). (G-I) ifa1-r ids1-mum1 кисточкой. (H) Пара колосков растения ifa1-r ids1-mum1 .Колосков образуется намного больше, чем у нормальных растений. (I) Внутри чешуек колоска формируется серия колосков. зр, колоск; зр *, лишний колоск; ф, цветочек; г, чешуйка; sm *, эктопическое соцветие, возникающее из СМ.
Уши двойного мутанта ifa1 ids1 обнаруживают фенотип, отличный от любого одного мутанта (рис. 8A-D). Внутри колосков возникает сильно разветвленная структура. Рассечение пары колосков (рис. 8E) и колосков (рис. 8F) выявляет основу сильно разветвленного фенотипа; внутри колосковых чешуек колоска образуются множественные соцветия и ветвистая структура (рис.8F). Эта разветвленная структура образует пары колосков в виде разветвленной структуры и, таким образом, считается ветвистой меристемой. Цветки на вторичной ветви повторяют цветочный фенотип ifa1-r . Поражаются оба колоска (рис. 8E).
Кисточки ifa1 ids1 двойных мутантов обнаруживают сильно разветвленный фенотип, при котором колоски образуются внутри колосков (Рис. 8G). На рис. 8I показан единственный колоск, содержащий колоски, выходящие из окружающих чешуек.Колоски, возникающие внутри колосков, обнаруживаются как на ножке, так и на сидячем колоске, а также в дополнительных колосках, которые образуются (Рис. 8H). Внематочные колоски развиваются поодиночке и последовательно, что позволяет предположить, что они возникают из меристемы пары колосков. Таким образом, ifa1 и ids1 действуют независимо для поддержания определенности SM и избыточно для поддержания его идентичности. Судьба СМ зависит от того, родится он на мужском или женском соцветии. У двойного мутанта SM уха становится BM, тогда как у кисточки SM становится SPM.
SEM использовали для отслеживания меристем, когда они инициируются у двойных мутантов (фиг. 9). Как показано на рис. 9В, мутанты ids1 и образуют лишние соцветия в колоске, которые возникают по типу двоякого ветвления. Дополнительный цветочек находится между верхним и нижним цветочками. У двойных мутантов ifa1 ids1 выпуклая меристема формируется между верхней и нижней меристемой цветков (Рис. 9C). Колоск справа (рис. 9C) немного более развит и показывает удлиненную меристему, которая, по-видимому, инициирует появление колосков.Более молодой колоск слева ясно показывает, что эта меристема возникает между двумя цветочками. СЭМ двойного мутанта ifa1 zag1 показывает аномальные верхнюю и нижнюю цветочные меристемы, которые инициируют несколько гребней (стрелки на фиг. 9D). Между верхним и нижним цветком меристемы не возникают. Эти результаты подчеркивают тот факт, что вторичные соцветия у мутантов ifa1 zag1 или ifa1 ids1 возникают из разных мест.
Рис. 9.
СЭМ развивающихся колосков.(A) Нормальная пара колосков. Более развитый верхний цветочек развивается на адаксиальной стороне, а нижний, частично покрытый внешней чешуей, формируется на абаксиальной стороне. (B) ids1-mum1 пара колосков. На одном из колосков образуется третий цветочек между верхним и нижним цветком. Гребень плодолистика (стрелка) дополнительного цветочка (f *) имеет ориентацию, противоположную ориентации верхнего цветочка и аналогичную ориентации нижнего цветочка. (C) Пара колосков из уха ifa1-r ids1-mum1 .Между двумя цветочками возникает неопределенная меристема (см *). Цветочные меристемы образуют дополнительные гребни плодолистиков, как видно у единственного мутанта ifa1 . (D) Продольный ряд из колосков ifa1-r zag1-mum1 . Боковые органы образуются из зачатка семяпочки (наконечники стрел). уф, верхний цветочек; lf — нижний цветочек; ог, наружная чешуя; ig, внутренняя чешуя; ул — тычинка; sm *, неопределенная меристема.
Мы также исследовали экспрессию zag1 и ids1 в мутантах ifa1 и обнаружили, что их паттерны экспрессии неотличимы от образцов дикого типа (данные не показаны).Эти результаты подтверждают генетический анализ, который не показал эпистатического взаимодействия.
ОБСУЖДЕНИЕ
Мы обнаружили у кукурузы ген, регулирующий детерминированность меристемы соцветия. Кукуруза имеет три типа детерминированных меристем: меристему пары колосков (SPM), меристему колосков (SM) и меристему цветков (FM). Мутация ifa1 наиболее серьезно влияет на цветочную меристему, но также влияет на две другие детерминированные меристемы.В одиночном мутанте ifa1 каждая детерминированная меристема продуцирует дополнительные последующие меристемы или органы, которые обычно определены. Однако в фоновом режиме, в котором мутирован либо zag1 , либо ids1 , каждая затронутая меристема продуцирует свои нормальные органы, а затем преобразуется в более ранние типы меристем. Какая меристема преобразуется и во что контролируется генетически. Преобразования различны без градиентов преобразования.
ifa1 требуется для определения цветочной меристемы
Цветочные меристемы дают четыре оборота органов у большинства цветков покрытосеменных, начиная с чашелистиков в качестве внешнего оборота, за которыми следуют лепестки, тычинки и, наконец, плодолистики.Плодолистики образуют семяпочки, содержащие женский гаметофит. Некоторые считают семяпочки пятым оборотом (Angenent et al., 1994; Ray et al., 1994). У кукурузы сросшиеся плодолистики составляют пестик, содержащий единственную семяпочку, являющуюся конечным продуктом цветочной меристемы (Sharman, 1945; Barnard, 1964). У мужских цветков пестик обрывается, а у женских цветков задерживается рост тычинок (Cheng et al., 1983; DeLong et al., 1993).
ifa1 мутанты кажутся нормальными на ранних стадиях развития цветков с правильно определенными органами цветков.Они начинают отличаться от дикого типа по мере того, как формируются плодолистики и зачаток семяпочки. Плодолистики не могут покрыть зачаток семяпочки, который продолжает расширяться. Расширяющийся зачаток семяпочки имеет много признаков меристемы, хотя он все еще экспрессирует маркер плодолистика / семяпочки, zag2 (Schmidt et al., 1993). Мужские цветки мутантов ifa1 также имеют дефект на верхушке цветка. У мутантов ifa1 в положении, обычно занимаемом прерванным пестиком, иногда обнаруживается выступающая нуцеллароподобная масса или дополнительные мужские соцветия.Частота неопределенности у мужских цветков не так высока, как у женских, но резко возрастает, когда пестик не прерывается. Такое состояние встречается у мутантов, таких как tasselseed2 (ts2) и Tasselseed5 ( Ts5 ), которые имеют феминизированные цветы на кисточке (Irish and Nelson, 1989; DeLong et al., 1993; Veit et al., 1993 ). У двойных мутантов ifa1 ts2 или ifa1 Ts5 все соцветия кисточки имеют выступающую нуцеллуподобную массу внутри тычинки (данные не представлены).Таким образом, неопределенный характер цветочной меристемы может преобладать над прерыванием пестика в мужском цветке и усиливается, когда прерывание пестика подавляется.
ifa1 требуется для определения идентичности колосковых и цветочных меристем
zag1 также регулирует детерминированность меристемы цветков (Mena et al., 1996). Сначала он экспрессируется в меристемах цветков, а затем в тычинках и плодолистиках, аналогично экспрессии AG в Arabidopsis (Yanofsky et al., 1990). Женские соцветия мутантов zag1 имеют неопределенную цветочную меристему, в которой образуются дополнительные плодолистики (Mena et al., 1996). Этот дефект аналогичен одному аспекту мутантов ifa1 . У двойных мутантов ifa1 zag1 цветочная меристема превращается в ветвящуюся меристему в женских цветках и превращается в колоск в мужских цветках после образования нормальных оборотов цветковых органов (рис. 10). Этот фенотип обнаруживает дополнительные роли как для zag1 , так и для ifa1 в женских и мужских цветках.Каждый ген функционирует независимо, чтобы поддерживать детерминированность женской цветочной меристемы, но функционирует избыточно, чтобы поддерживать свою идентичность. Хотя zag1 не играет очевидной роли в мужских цветках, как zag1 , так и ifa1 действуют избыточно, чтобы поддерживать идентичность цветочной меристемы в мужских цветках.
Рис. 10.
Идентичность меристем у разных мутантов. IM — меристема соцветия; BM — меристема ветви; SPM — меристема пары колосков; SM — меристема колосков; FM — цветочная меристема; FA, цветочная вершина; тонкая стрелка, меристема определенная; толстая стрелка, неопределенная меристема, дающая дополнительные органы или меристемы.
Пенетрантность фенотипа большого нуцеллуса у одиночных мутантов ifa1 составляет 100%, как и пенетрантность фенотипа ветвящейся меристемы у двойных мутантов ifa1 zag1 . Иногда одно ядро на ухе мутанта ifa1 может проявлять фенотип ветвления. Хотя это открытие может указывать на то, что ifa1 сам по себе может регулировать идентичность, также возможно, что zag1 или гены, расположенные ниже zag1 , не могут функционировать с некоторой стохастической частотой.
Синергетическое взаимодействие также происходит у двойных мутантов ifa1 ids1 . ids1 одиночных мутантов имеют дополнительные соцветия как в мужских, так и в женских колосках (Chuck et al., 1998). ids1 экспрессируется в незрелых органах цветка, паре колосков и меристемах колосков. У двойного мутанта ifa1 ids1 SM в ухе продуцирует цветочные меристемы, как обычно, но затем превращается в меристему ветвления. SM в кисточке также преобразуется после образования цветочных меристем.Эта мужская меристема дает серию колосков, которые инициируются по отдельности и, таким образом, могут рассматриваться как ВП.
Несмотря на эффект ifa1-r на все детерминированные меристемы, ids1-mum1 и zag1-mum1 ограничивают превращение, наблюдаемое в двойных мутантах, до одного типа меристемы. ids1 играет роль в детерминированности SM, и, таким образом, двойной мутант влияет на конверсию SM. zag1 играет роль в детерминации цветочной меристемы, и, таким образом, двойной мутант влияет на конверсию цветочной меристемы (рис.10). Независимо от того, превращаются ли меристемы двойных мутантов в ветвь, пару колосков или колосков, реверсии являются полными, а не смесью типов меристем. Этот результат предполагает, что идентичность каждого типа меристемы четко определена. Это открытие интригует, учитывая, что ifa1 влияет на все меристемы и ids1 экспрессируется на всех стадиях, но их двойное отсутствие влияет на один тип меристемы наиболее значительно. Это предполагает, что некоторые другие неидентифицированные гены должны функционировать только на стадии SM.
Превращения самцов и самок различны у обоих двойных мутантов (рис. 10). В каждом случае преобразующая женская меристема становится меристемой ветвления, а преобразующая мужская меристема становится меристемой парных колосков или колосков. Мы также обнаружили, что степень выраженности фенотипа ifa1-r различалась у мужских цветков, которые были феминизированы или нет. Другие мутанты кукурузы также более опасны для уха, чем кисточка. Например, уши мутанта bd1 имеют SM, которые продуцируют BM, и никогда не достигают стадии цветочной меристемы. bd1 мутантные кисточки , с другой стороны, способны продуцировать цветочные меристемы (G. Chuck, S.H. и R. Schmidt, неопубликовано) (Colombo et al., 1998). Разница между мужским и женским соцветиями может быть результатом разного уровня гормонов или различной чувствительности к гормонам из-за разного положения на растении (Dellaporta and Calderon-Urrea, 1994; Irish, 1996).
Конверсия Meristem
Классический пример преобразования меристемы обнаружен в Impatiens balsamina (Battey and Lyndon, 1990). Impatiens требует постоянных коротких дней для цветения. Если растения возвращаются из индуктивных коротких дней в длинные, цветочная меристема начинает давать вегетативные листья из цветочной меристемы. Иногда образуются химерные органы, состоящие частично из лепестка и частично из листа. Возврат к короткому дню снова приводит к немедленному производству лепестков. Возврат к вегетативному развитию также наблюдается в мутации undeterminate1 ( id1 ) кукурузы (Colasanti et al., 1998). id1 мутанты не могут зацвести в большинстве нормальных сезонов роста и дают в два-три раза больше листьев, чем обычно. В конце концов, апикальная меристема побега превращается в кисточку, которая в высшей степени аномальна. На этих растениях не образуются уши. Рассечение кисточки id1 показывает, что вегетативные проростки инициируются из меристемы колосков после зарождения двух цветков (Galinat and Naylor, 1951) (D. L.C. и J. Colasanti, неопубликовано). Таким образом, меристема колоска id1 превратилась в вегетативную.
Ряд мутаций влияет на детерминированность и идентичность меристемы цветка Arabidopsis . AGAMOUS ( AG ) играет решающую роль в детерминации цветочной меристемы у Arabidopsis (Bowman et al., 1989). При определенных условиях цветочная меристема ag дает меристему соцветия в центре мутанта плодолистика, аналогичную двойным мутантам ifa1 zag1 . Окамура и его коллеги показали, что агамных гомозигот или листовых ( lfy ) гетерозигот в условиях короткого дня дают побеги изнутри мутовки плодолистика (Okamuro et al., 1993). Аналогичные результаты были получены при использовании двойных мутантов ag constans (Mizukami and Ma, 1997). CONSTANS регулирует LFY (Simon et al., 1996), который сам является активатором AG (Busch et al., 1999), что объясняет, почему ограничение количества LFY может приводить к тому же фенотипу. Условия короткого дня задерживают активацию LFY (Busch et al., 1999) и поэтому, возможно, также уменьшают количество AG.
Недавно две группы показали, что LFY вместе с WUSCHEL, гомеодоменным белком, который необходим для функции меристемы (Lenhard et al., 2001; Lohmann et al., 2001), положительно регулируют AG и что AG отрицательно регулирует WUS. Экспрессия WUS сохраняется в неопределенных меристемах ag цветков (Lohmann et al., 2001). Мы показали, что kn1 экспрессируется в зачатках семяпочек мутантов ifa1 , что позволяет предположить, что IFA1 негативно регулирует KN1. Учитывая параллели между ifa1 zag1 и слабыми ag аллелями Arabidopsis , вполне вероятно, что IFA1 также негативно регулирует ортолог WUS.
Модели для архитектуры соцветий кукурузы
Различные модели рассматривали взаимоотношения меристем ветвей кукурузы. Модель конверсии предполагает, что SPM превращается в SM после инициирования сидячего колоска, а SM преобразуется в верхнюю меристему цветка после инициирования нижнего цветочка (Irish, 1997). С помощью этой модели фенотип ifa1-r можно объяснить задержкой превращения одного типа меристемы в другой.Эта задержка привела бы к инициированию большего количества меристем перед конверсией. SPM инициирует более одного SM, а SM инициирует более одной цветочной меристемы перед преобразованием. Возможно, если SM может преобразоваться в цветочную меристему, то он также может преобразоваться обратно в SPM, в зависимости от количества генного продукта. Другая модель предполагает, что остаточная SM присутствует после продуцирования обеих цветочных меристем (Chuck et al., 1998; Irish, 1998). У мутантов, таких как ids1-mum1 или ifa1-r , остаточная меристема продолжает продуцировать боковые органы.Наш SEM-анализ ifa1 ids1 подтверждает модель остаточной меристемы, в которой меристема явно присутствует между двумя цветочными меристемами.
Заключение
Колоски объединяют все травы, однако ветви, на которых рождаются колоски, значительно различаются в зависимости от вида (Clifford, 1987). Колоски могут возникать непосредственно на первичном соцветии или на вторичных или третичных ветвях, и они могут давать один, два или несколько соцветий.У Andropogoneae, подгруппы трав, в которую входит кукуруза, есть два цветочка на колоске и два колоска на модифицированной ветви, называемой парой колосков. Уникальное строение Andropogoneae считается свидетельством монофилии подсемейства (Kellogg, Campbell, 1987). ifa1 и несколько других генов ответственны за детерминированность SPM и SM. Интересно предположить, был ли ifa1 рекрутирован из цветочных меристем в меристемы ветвления и, таким образом, ответственен за детерминированность SPM и SM.Исследование экспрессии и функции генов детерминированности меристемы ветвей у разных трав может ответить на эти вопросы и привести к пониманию эволюции морфологии травы.
Благодарности
Мы благодарим сотрудников лаборатории Хека за помощь в этой области, интересные обсуждения и вклад в эту статью, Р. Шмидта за клон кДНК zag2 , Трейси Ямаваки за картирование ifa1-r относительно zmm14 и Е. .Фоллбрехту за предоставление аллеля ifa1 из его поля Mu . Особая благодарность Джорджу Чаку, Нику Каплински, Юки Мизуками, Леоноре Райзер и Марку Бегу за критическое прочтение рукописи. Мы благодарны Дэвиду Ханцу за заботу о наших растениях в теплице и Мануэлю Айзенбергу за помощь в скрининге полей Mu на ifa1 аллелей. Эта работа была поддержана Министерством сельского хозяйства США, CRIS 5335-21000-018-00D.D. L.C. был поддержан докторской стипендией NSF (Премия № 28).
Каталожные номера
- ↵
Амвросий Б.А., Лернер Д.Р., Цицери П., Падилья К.М., Янофски М.Ф. и Шмидт Р.Дж. (2000). Молекулярный и генетический анализ гена silky1 показывает сохранение в спецификации органов цветка у эвдикотов и однодольных. Мол. Cell 5, 569-579.
- ↵
Ангенент, Г. К., Франкен, Дж., Busscher, M., Weiss, D. и Vantunen, A.J. (1994). Совместное подавление гомеотического гена петунии fbp2 влияет на идентичность генеративной меристемы. Завод J . 5, 33-44.
- ↵
Барнард, К. (1964). Форма и структура. В Grasses and Grasslands (ed. C. Barnard), стр. 47-72. Лондон: Макмиллан.
- ↵
Бэтти, Н. Х. и Линдон, Р. Ф. (1990).Возврат цветения. Бот. Ред. . 56, 162–189.
- ↵
Beckett, J. B. (1993). Размещение рецессивных генов в плече хромосомы с транслокациями B-A. В The Maize Handbook (ред. М. Фрилинг и В. Уолбот), стр. 315-327. Нью-Йорк, штат Нью-Йорк: Springer Verlag.
- ↵
Bowman, J. L., Smyth, D. R. и Meyerowitz, E. M. (1989). Гены, управляющие развитием цветка у Arabidopsis.Растительная клетка 1, 37-52.
- ↵
Брэдли Д., Карпентер Р., Соммер Х., Хартли Н. и Коэн Е. (1993). Дополнительные цветочные гомеотические фенотипы являются результатом противоположной ориентации транспозона в локусе plena Antirrhinum. Ячейка 72, 85-95.
- ↵
Буш М. А., Бомблис К. и Вейгель Д. (1999). Активация цветочного гомеотического гена у Arabidopsis. Science 285, 585-587.
- ↵
Cheng, P. C., Greyson, R. I. and Walden, D. B. (1983). Зарождение органа и развитие однополых цветков в кисточке и колосе Zea mays . Являюсь. Дж. Бот . 70, 450-462.
- ↵
Чак Г., Мили Р. и Хейк С. (1998). Контроль судьбы меристемы колосков кукурузы с помощью APETALA2-подобного гена неопределенного spikelet1. Гены Дев . 12, 1145-1154.
- ↵
Клиффорд, Х. Т. (1987). Морфология колосков и цветков. In Grass Systematics and Evolution (ред. Т. Р. Содерстром, К. В. Хилу, К. С. Кэмпбелл и М. Э. Баркворт), стр. 21-30. Вашингтон, округ Колумбия: Пресса Смитсоновского института.
- ↵
Colasanti, J., Yuan, Z. and Sundaresan, V. (1998). Неопределенный ген кодирует белок цинкового пальца и регулирует генерируемый листьями сигнал, необходимый для перехода к цветению кукурузы.Cell 93, 593-603.
- ↵
Коломбо, Л., Марциани, Г., Мазиеро, С., Виттих, П. Э., Шмидт, Р. Дж., Горла, М. С. и Пе, Э. М. (1998). Ветвистая бесшелковая опосредует переход от колосковой меристемы к цветочной во время развития колоса Zea mays. Завод J . 16, 353-363.
- ↵
Dellaporta, S. L. и Calderon-Urrea, A. (1994). Процесс определения пола кукурузы.Science 266, 1501–1505.
- ↵
Делонг А., Кальдерон-Урреа А. и Деллапорта С. Л. (1993). Ген детерминации пола TASSELSEED2 кукурузы кодирует короткоцепочечную алкогольдегидрогеназу, необходимую для специфического аборта цветочного органа. Cell 74, 757-768.
- ↵
Galinat, W. C. и Naylor, A. W. (1951). Связь фотопериода с размножением соцветий у Zea mays L.Являюсь. Дж. Бот . 38, 38-47.
- ↵
Irish, E. E. (1996). Регуляция определения пола у кукурузы. BioEssays 18, 363-369.
- ↵
Irish, E. E. (1997). Мутации семян метелки класса II свидетельствуют о множестве типов меристем соцветий у кукурузы (Poaceae). Являюсь. Дж. Бот . 84, 1502-1515.
- ↵
Айриш, Э.E. (1998). Колоски травы: колючая проблема. BioEssays 20, 789-793.
- ↵
Айриш, Э. Э. и Нельсон, Т. (1989). Определение пола у однодомных и двудомных растений. Растительная клетка 1, 737-744.
- ↵
Джексон Д., Вейт Б. и Хейк С. (1994). Экспрессия генов гомеобокса, родственных кукурузе KNOTTED1, в апикальной меристеме побега предсказывает паттерны морфогенеза вегетативного побега.Развитие 120, 405-413.
- ↵
Kellogg, E. A. и Campbell, C. S. (1987). Филогенетический анализ злаков. In Grass Systematics and Evolution (ред. Т. Р. Содерстром, К. В. Хилу, К. С. Кэмпбелл и М. Э. Баркворт), стр. 310-322. Вашингтон, округ Колумбия: Пресса Смитсоновского института.
- ↵
Кемптон, Дж. Х. (1934). Унаследованные признаки кукурузы: XLVII Ветвистые, без шелка.Дж. Херед . 25, 29-32.
- ↵
Kerstetter, R.A., Laudencia-Chingcuanco, D., Smith, L.G. и Hake, S. (1997). Мутации с потерей функции в гене гомеобокса кукурузы, knotted1, нарушают поддержание меристемы побегов. Разработка 124, 3045-3054.
- ↵
Laughnan, J. R. и Gabay-Laughnan, S. (1993). Размещение генов с помощью реципрокных транслокаций с восковой меткой.В The Maize Handbook (ред. М. Фрилинг и В. Уолбот), стр. 254-257. Нью-Йорк: Springer Verlag.
- ↵
Lenhard, M., Bohnert, A., Jurgens, G. and Laux, T. (2001). Прекращение поддержания стволовых клеток в цветочных меристемах Arabidopsis за счет взаимодействий между WUSCHEL и AGAMOUS. Ячейка 105, 805-814.
- ↵
Ломанн, Дж. У., Хонг, Р. Л., Хобе, М., Буш, М. А., Парси, Ф., Саймон Р. и Вейгель Д. (2001). Молекулярная связь между регуляцией стволовых клеток и формированием цветочного узора у Arabidopsis. Ячейка 105, 793-803.
- ↵
Макстин П., Лауденсия-Чингкуанко Д. и Коласанти Дж. (2000). Цветочек под любым другим названием: контроль идентичности меристемы кукурузы. Тенденции Завод Sci . 5, 61-66.
- ↵
Мена, М., Амброуз, Б., Мили, Р. Б., Бриггс, С.П., Янофски М. Ф. и Шмидт Р. Дж. (1996). Диверсификация активности С-функции в развитии цветков кукурузы. Наука 274, 1537-1540.
- ↵
Mizukami, Y. and Ma, H. (1997). Определение идентичности цветочной меристемы арабидопсиса методом AGAMOUS. Растительная клетка 9, 393-408.
- ↵
Никерсон, Н. Х. (1954). Морфологический анализ початка кукурузы. Являюсь. Дж. Бот . 41, 87-92.
- ↵
Окамуро, Дж. К., ден Бур, Б. Г. У. и Джофуку, К. Д. (1993). Регуляция развития цветка арабидопсиса. Растительная клетка 5, 1183-1193.
- ↵
Постлтуэйт, С. Н. и Нельсон, О. Е. (1964). Характеристика развития кукурузы с использованием мутантов I. Политипические (Pt) и ramosa-1 (ra1) мутанты. Являюсь. Дж. Бот . 51, 238-243.
- ↵
Рэй, А., Робинсон-Бирс, К., Рэй, С., Бейкер, С. К., Ланг, Дж. Д., Пройс, Д., Миллиган, С. Б. и Гассер, С. С. (1994). Гомеотический ген белл (bel1) арабидопсиса контролирует развитие семяпочки посредством негативной регуляции гена AGAMOUS (AG). Proc. Natl. Акад. Sci. USA 91, 5761-5765.
- ↵
Schmidt, R.J., Veit, B., Mandel, M.A., Mena, M., Hake, S. и Yanofsky, M. F. (1993). Идентификация и молекулярная характеристика ZAG1, гомолога кукурузы цветочного гомеотического гена AGAMOUS Arabidopsis.Растительная клетка 5, 729-737.
- ↵
Шарман Б.С. (1945). Зарождение листьев и почек у злаковых. Бот. Газ . 106, 269-289.
- ↵
Саймон Р., Игено М. И. и Коупленд Г. (1996). Активация генов идентичности цветочной меристемы у Arabidopsis. Nature 384, 59-62.
- ↵
Смит, Л. Г., Грин, Б., Вейт, Б. и Хейк, С. (1992). Доминантная мутация в гене гомеобокса кукурузы, Knotted-1 , вызывает его эктопическую экспрессию в клетках листа с измененной судьбой. Развитие 116, 21-30.
- ↵
Steeves, T. A. and Sussex, I. M. (1989). Образцы в развитии растений 2-е изд. Кембридж: Издательство Кембриджского университета.
- ↵
Вейт Б., Шмидт Р. Дж., Хейк С. и Янофски М. Ф. (1993). Цветочное развитие кукурузы — новые гены и старые мутанты. Растительная клетка 5, 1205-1215.
- ↵
Yanofsky, M. F., Ma, H., Bowman, J. L., Drews, G. N., Feldmann, K. A. и Meyerowitz, E. M. (1990). Белок, кодируемый гомеотическим геном Arabidopsis agamous, напоминает факторы транскрипции. Nature 346, 35-39.
подавитель сидячих колосков1 Функции в пути ветвления, контролирующие детерминантность меристем у кукурузы | Физиология растений
Абстрактные
Колосок, представляющий собой короткую ветвь, несущую соцветия, является фундаментальной единицей архитектуры соцветий травы.У большинства трав колоски поодиночке расположены на соцветиях. Однако парные колоски характерны для Andropogoneae, трибы из 1000 видов, включая кукурузу ( Zea mays ). Супрессор сидячих колосков1 ( Sos1 ) мутант кукурузы дает одиночные, а не парные колоски в соцветии. Следовательно, ген sos1 мог участвовать в эволюции парных колосков. В этой статье мы показываем, что Sos1 является полудоминантной антиморфной мутацией. Sos1 Мутанты имеют меньше ветвей и колосков по двум причинам: (1) образуется меньше меристем пар колосков из-за дефектов размера меристемы соцветий и (2) образующиеся меристемы пары колосков образуют одну вместо двух меристем колосков. Было исследовано взаимодействие Sos1 с мутантами ramosa , которые производят больше ветвей и колосков. Результаты показывают, что Sos1 имеет эпистатическое взаимодействие с ramosa1 ( ra1 ), синергетическое взаимодействие с ra2 и аддитивное взаимодействие с ra3 .Более того, уровни мРНК ra1 снижены у мутантов Sos1 , в то время как уровни мРНК ra2 и ra3 остаются неизменными. Основываясь на этих генетических исследованиях и исследованиях экспрессии, мы предполагаем, что sos1 функционирует в ветви ra1 пути ramosa , контролируя детерминированность меристемы.
Органогенез растений контролируется меристемами (Steeves and Sussex, 1989). Органы образуются в периферической зоне меристемы, в то время как в центральной зоне сохраняется пул клеток, которые не дифференцируются, что позволяет меристеме поддерживать себя.Меристемы определяются по их детерминированности, идентичности и положению (McSteen et al., 2000). Детерминантные меристемы продуцируют ограниченное количество структур перед окончанием, тогда как неопределенные меристемы обладают потенциалом продолжать продуцировать органы неопределенно долго (Bortiri and Hake, 2007; Sablowski, 2007). Апикальные меристемы у многих видов индетерминантные. Апикальная меристема побега (САМ) дает начало вегетативному побегу, а апикальная меристема соцветия дает начало цветущему побегу.Пазушные меристемы, образующиеся в пазухах зачатков листьев, могут быть неопределенными и давать ветви или могут быть детерминантными и давать начало цветкам.
В развитии соцветий кукурузы ( Zea mays ) существует несколько типов пазушных меристем, которые различаются по своей детерминированности, что приводит к сильно разветвленному соцветию (Irish, 1997a; Bortiri and Hake, 2007; Barazesh and McSteen, 2008b). Созревшее мужское соцветие состоит из главного колоса и нескольких длинных боковых ветвей, которые прикрыты короткими ветвями, называемыми парами колосков (рис.1A
Рис. 1.
Sos1 фенотип кисточки и ушка. A, фотография зрелых кисточек до цветения. Обычные кисточки имеют длинные ветки у основания главного шипа. Пары колосков покрывают ветви и главный колос. Мутанты Sos1 / + имеют редкий вид из-за образования меньшего количества ветвей и колосков. Sos1 / Sos1 Мутанты имеют очень мало ветвей и одиночных колосков. B, количественное определение количества веток в кисточке. C. Количественная оценка количества парных (серая полоса) по сравнению с одиночными (белыми) колосками на главном колоске кисточки.D. Фотография зрелых колосьев после открытого опыления. Нормальные уши имеют парные ряды ядер. Уши Sos1 / + и Sos1 / Sos1 имеют меньшее количество рядов ядер в ухе. E, Количественная оценка количества ядер на ухо. F, Количественное определение количества рядов ядер на ухо. Sos1 / Sos1 уши имеют менее половины количества рядов по сравнению с обычными братьями и сестрами. Все графики являются средними ± стандартными значениями. [Цветную версию этого рисунка см. В статье в Интернете.]
Рисунок 1.
Sos1 фенотип кисточки и ушка. A, фотография зрелых кисточек до цветения. Обычные кисточки имеют длинные ветки у основания главного шипа. Пары колосков покрывают ветви и главный колос. Мутанты Sos1 / + имеют редкий вид из-за образования меньшего количества ветвей и колосков. Sos1 / Sos1 Мутанты имеют очень мало ветвей и одиночных колосков. B, количественное определение количества веток в кисточке. C. Количественная оценка количества парных (серая полоса) по сравнению с одиночными (белыми) колосками на главном колоске кисточки.D. Фотография зрелых колосьев после открытого опыления. Нормальные уши имеют парные ряды ядер. Уши Sos1 / + и Sos1 / Sos1 имеют меньшее количество рядов ядер в ухе. E, Количественная оценка количества ядер на ухо. F, Количественное определение количества рядов ядер на ухо. Sos1 / Sos1 уши имеют менее половины количества рядов по сравнению с обычными братьями и сестрами. Все графики являются средними ± стандартными значениями. [Цветную версию этого рисунка см. В статье в Интернете.]
; Киссельбах, 1949).Колоск определяется как короткая ветвь с двумя листовидными чешуйками, окружающими соцветия (Clifford, 1987). Во время развития соцветия меристема верхушечного соцветия (IM) производит пазушные меристемы, называемые меристемами ветвей (BM), которые являются неопределенными и производят ветви. После того, как сделано несколько ветвей, IM резко переключается на производство меристем пар колосков (SPM), которые являются детерминированными, поскольку они производят два колоска. Меристемы колосков (SM) также детерминированы, поскольку они производят две цветочные меристемы (FM), которые затем производят цветочные органы.Хотя у кукурузы есть отдельные мужские и женские соцветия, называемые кисточкой и колосом соответственно, раннее развитие похоже, за исключением того, что колосья не дает ветвей.
Были предложены две модели, чтобы объяснить, как SPM производит два SM, а SM производит два FM (Irish, 1997a, 1998; Chuck et al., 1998; Kaplinsky and Freeling, 2003). В модели латерального ветвления SPM продуцирует две SM посредством бокового ветвления, а остаточная меристема остается между двумя SM (Chuck et al., 1998). В модели преобразования SPM инициирует SM, а затем оставшийся SPM преобразуется в SM (Irish, 1997a, 1998).Сильную поддержку модели конверсии недавно предоставили эксперименты, в которых нормальные растения кукурузы обрабатывали ингибиторами транспорта ауксина, которые ингибировали инициацию SM, но не ингибировали превращение SPM в SM, что приводило к образованию одиночных колосков (Wu and McSteen , 2007).
Супрессор сидячих колосков1 мутант ( Sos1 ) производит одиночные, а не парные колоски в соцветии (Doebley et al., 1995). У нормальных растений один колосок прикреплен к позвоночнику соцветия ножкой и называется колоском на ножке, а другой колоск, не имеющий цветоножки, называется сидячим колоском.У мутантов Sos1 сидячий колоск не образуется, что приводит к образованию единичных колосков на ножке. Следовательно, у мутантов Sos1 SPM продуцирует один SM вместо двух и, следовательно, является более детерминированным, чем обычно.
Детерминированность SPM положительно регулируется путем ramosa (McSteen, 2006; Kellogg, 2007). ramosa1 ( ra1 ) кодирует фактор транскрипции цинкового пальца, экспрессируемый в основании SPM (Vollbrecht et al., 2005). У мутантов ra1 кисточка и початок сильно разветвлены, потому что SPM становятся неопределенными и вырастают, давая дополнительные колоски (Gernart, 1912; Vollbrecht et al., 2005). ra1 действует ниже ra2 , который кодирует фактор транскрипции LOB-домена, экспрессируемый в зачатке BM, SPM и SM (Bortiri et al., 2006a). ra1 также действует после ra3 , который кодирует трегалозо-6-фосфатфосфатазу (Satoh-Nagasawa et al., 2006). Интересно, что хотя ra2 и ra3 -подобные гены присутствуют в других злаках, таких как рис ( Oryza sativa ), ra1 был идентифицирован только у Andropogoneae, клады, к которой принадлежит кукуруза.
В подавляющем большинстве злаков, включая рис, ячмень ( Hordeum vulgare ) и пшеницу ( Triticum aestivum ), колоски образуются поодиночке (Clifford, 1987). Производство парных колосков характерно для Andropogoneae, племени из 1000 видов, включая кукурузу, мискантус , сахарный тростник ( Saccharum officinarum ) и сорго ( Sorghum bicolor ; Kellogg, 2000, 2001; Grass Phylogeny Working Group, 2001).В племени Paniceae, которое является сестрой Andropogoneae, у большинства видов есть одиночные колоски, но некоторые виды производят парные колоски, а другие производят колоски, соединенные с щетинками, которые представляют собой модифицированные колоски (Kellogg, 2000; Doust and Kellogg, 2002). . Следовательно, парный колоск является производным признаком, хотя неясно, когда этот признак возник во время эволюции Panicoideae. Было высказано предположение, что ra1 могли быть задействованы для эволюции признака парных колосков (Vollbrecht et al., 2005; Макстин, 2006). Поскольку мутанты Sos1 продуцируют одиночные, а не парные колоски, ген sos1 также может быть вовлечен в эволюцию парных колосков.
В этой статье мы используем количественный анализ, сканирующую электронную микроскопию (SEM) и гистологию, чтобы показать, что мутация Sos1 вызывает дефекты в развитии IM, BM и SPM как в кисточке, так и в ухе. Мы используем анализ дозировки, чтобы показать, что Sos1 является антиморфом, то есть доминантно-отрицательной мутацией.Более того, генетический анализ и анализ экспрессии предоставляют доказательства того, что ген sos1 действует в ветви ra1 пути ramosa , контролируя детерминированность меристемы.
РЕЗУЛЬТАТЫ
Аллель Sos1-Reference возник спонтанно (Doebley et al., 1995). Семена были получены из кукурузного генетического кооператива и подвергнуты обратному скрещиванию с получением B73. Ранее сообщалось, что мутант отображается на коротком плече хромосомы 4 (Doebley et al., 1995). Для создания картирующей популяции Sos1 (на фоне B73) скрещивали с генетическим фоном Mo17 и подвергали обратному скрещиванию с Mo17. Маркеры простых повторов последовательности (SSR) использовали для дальнейшего определения местоположения Sos1 с точностью до 4 сМ от umc1294 в ячейке 4.02. Поскольку маркер umc1294 был полиморфен между B73 и исходным фоном, на котором возник Sos1 , этот маркер использовали для генотипирования Sos1 на фоне B73.Генотипирование в сочетании с анализом фенотипа определили, что мутация Sos1 была полудоминантной, а не доминантной, как сообщалось ранее (Рис. 1; Doebley et al., 1995).
Sos1 Мутанты производят меньше ветвей и колосков
Мутанты Sos1 имеют дефекты кисточки и ушка. В метелке мутантных растений Sos1 давали меньше ветвей и колосков (рис. 1, A – C). Семьи, сегрегированные по Sos1 , были генотипированы, и количество ветвей и колосков было подсчитано у нормальных братьев и сестер по сравнению с растениями, гетерозиготными и гомозиготными по Sos1 .Эти результаты показали, что мутанты Sos1 / + и Sos1 / Sos1 давали меньше ветвей, чем обычно (Рис. 1B). Для анализа дефектов колосков отдельно подсчитывали количество парных колосков по сравнению с одиночными. В то время как у нормальных растений были парные колоски, у мутантов Sos1 / + и Sos1 / Sos1 было больше одиночных, чем парных колосков (рис. 1С).
Мутация Sos1 также затронула ухо. Нормальные уши не разветвлены, но дают пары колосков (Kiesselbach, 1949).Каждый колоск дает один цветочек (нижний цветочек обрывается), который при опылении дает одно ядро (Cheng et al., 1983). Колоски в колосе сидячие, поэтому спаривание не очевидно, но оно приводит к четному количеству рядов ядер в зрелом колосе. У мутантов Sos1 в ухе было меньше ядер (рис. 1D). Количественная оценка показала, что общее количество ядер было уменьшено у мутантов Sos1 / + и Sos1 / Sos1 по сравнению с нормальными ушами (рис.1E). Также наблюдалось уменьшение количества рядов ядер с мутантами Sos1 / + , дающими переменное количество рядов, и мутантами Sos1 , дающими менее половины количества рядов нормальных братьев и сестер (рис. 1F).
Чтобы определить основу развития для производства меньшего количества ветвей и колосков у мутантов Sos1 , СЭМ-анализ был выполнен на развивающихся кистях и ушах. В нормальных соцветиях ВП производятся из ИМ на верхушке соцветия (рис.2А
Рис. 2. СЭМ
соцветий Sos1 . От А до С, СЭМ ушей 8-недельного возраста. A, Нормальное ухо производит ряды SPM на кончике уха, которые переходят в производство парных SM ниже кончика. B, Sos1 / + ухо, производящее одиночный или парный SM. C, Sos1 / Sos1 ухо производящее одиночное SM. От D до F, большее увеличение SM в ухе. D, Нормальное ухо с парным СМ. Наружная чешуйка (og) и внутренняя чешуя (ig) являются первыми органами, производимыми SM.E, Sos1 / + ухо, производящее одиночный или парный SM. F, Sos1 / Sos1 ухо с одинарным SM. G, обычная кисточка, создающая парные SM. H, Sos1 / + кисточка с парным SM вверху и одиночным SM у основания. I, Sos1 / Sos1 кисточкой, производящей ряды одиночных SM. Масштабная линейка = 100 мкм м от A до C и от G до I и 50 мкм м от D до F.
Рисунок 2.
СЭМ соцветий Sos1 . От А до С, СЭМ ушей 8-недельного возраста.A, Нормальное ухо производит ряды SPM на кончике уха, которые переходят в производство парных SM ниже кончика. B, Sos1 / + ухо, производящее одиночный или парный SM. C, Sos1 / Sos1 ухо производящее одиночное SM. От D до F, большее увеличение SM в ухе. D, Нормальное ухо с парным СМ. Наружная чешуйка (og) и внутренняя чешуя (ig) являются первыми органами, производимыми SM. E, Sos1 / + ухо, производящее одиночный или парный SM. F, Sos1 / Sos1 ухо с одинарным SM.G, обычная кисточка, создающая парные SM. H, Sos1 / + кисточка с парным SM вверху и одиночным SM у основания. I, Sos1 / Sos1 кисточкой, производящей ряды одиночных SM. Масштабная линейка = 100 мкм м от A до C и от G до I и 50 мкм м от D до F.
). Затем SPM производит два SM (рис. 2, A и D). На идентичность меристемы как на SM указывает образование чешуек, защитных листоподобных органов, которые являются первыми органами, продуцируемыми SM (Cheng et al., 1983). Наружная чешуйка образуется на абаксиальной стороне SM, а внутренняя чешуйка формируется внутри внешней чешуи на адаксиальной стороне SM (рис. 2D). Во время развития уха мутанты Sos1 / + и Sos1 / Sos1 продуцировали SPM как обычно, за исключением того, что оказалось меньше рядов (рис. 2, B и C). Позже в развитии уха, когда парные SM были видны в нормальных ушах, у мутантов Sos / Sos1 и были одиночные SM, а у мутантов Sos1 / + была смесь одиночных и парных SM (рис.2, Г – Ж). Аналогично, в метелке SPM нормальных растений продуцировали два SM (рис. 2G), в то время как мутанты Sos / Sos1 продуцировали одиночные SM (рис. 2I), а мутанты Sos1 / + продуцировали один или два SM (рис. 2H). Следовательно, у мутантов Sos1 и SPM продуцируют один SM вместо двух.
Sos1 Мутанты имеют дефекты размера IM
Если бы единственным дефектом у мутантов Sos1 было образование одиночных, а не парных колосков, то можно было бы ожидать, что мутанты производят ровно половину нормального количества рядов ядра.Однако мутанты Sos1 / Sos1 продуцировали менее половины нормального числа рядов ядер (Fig. 1F; Doebley et al., 1995). Более того, мутанты Sos1 / Sos1 продуцировали менее половины нормального количества колосков в кисточке (рис. 1С). Это указывает на дополнительный дефект в инициировании SPM. Для количественной оценки дефекта в инициировании SPM был использован анализ SEM для подсчета количества SPM, инициированных по окружности IM. Это показало, что нормальные уши производили 9 или 10 SPM, Sos1 / + давали в среднем 7.37 ± 0,33 SPM, в то время как Sos1 / Sos1 продуцировал в среднем 6,14 ± 0,34 SPM (Рис. 3A
Рис. 3.
Анализ IM мутантов Sos1 . A, Количественная оценка количества SPM, продуцируемых около окружность кончика ушей Sos1 . B, Измерение ширины IM. C, Измерение высоты IM. D до F, SEM IM нормального, Sos1 / + и Sos1 / Sos1 уши. Масштаб = 50 мкм м.
Рисунок 3.
Анализ IM мутантов Sos1 . A, Количественное определение количества SPM, произведенных по окружности кончика ушей Sos1 . B, Измерение ширины IM. C, Измерение высоты IM. От D до F, SEM IM нормальных, Sos1 / + и Sos1 / Sos1 ушей. Масштабная линейка = 50 мкм м.
). Для дальнейшего анализа дефектов инициации SPM соцветия заливали воском, делали срезы и окрашивали толуидиновым синим О (ТВО).Около кончика уха были видны СЗМ кольцом по окружности соцветия. На срезах, показанных на рисунке 4
Рисунок 4.
Поперечные сечения ушей Sos1 , окрашенные ТВО. От А до С: поперечные сечения ушей около кончика уха на стадии производства СЗМ. A, Нормальное ухо показывает девять SPM, некоторые из которых только начинают формировать SM. B, Sos1 / + ухо, показывающее восемь SPM. C, Sos1 / Sos1 уши, показывающие шесть SPM. От D до F, поперечные сечения позже в развитии, когда нормальные уши дали парные SM.D — Нормальное ухо с девятью парами SM. Обратите внимание, что внешняя чешуйка (g) видна на некоторых SM. E, Sos1 / + ухо, в котором два из восьми SPM производят два SM. F, Sos1 / Sos1 ухо, в котором видны шесть одиночных SM. Все SM производят чешуйки (g). Масштабная линейка = 100 мкм м. [Цветную версию этого рисунка см. В статье в Интернете.]
Рис. 4.
Поперечные сечения ушей Sos1 , окрашенные ТВО. От А до С: поперечные сечения ушей около кончика уха на стадии производства СЗМ.A, Нормальное ухо показывает девять SPM, некоторые из которых только начинают формировать SM. B, Sos1 / + ухо, показывающее восемь SPM. C, Sos1 / Sos1 уши, показывающие шесть SPM. От D до F, поперечные сечения позже в развитии, когда нормальные уши дали парные SM. D — Нормальное ухо с девятью парами SM. Обратите внимание, что внешняя чешуйка (g) видна на некоторых SM. E, Sos1 / + ухо, в котором два из восьми SPM производят два SM. F, Sos1 / Sos1 ухо, в котором видны шесть одиночных SM.Все SM производят чешуйки (g). Масштабная линейка = 100 мкм м. [Цветную версию этого рисунка см. В онлайн-статье.]
, нормальное ухо инициировало девять SPM (фиг. 4A), гетерозиготное ухо инициировало восемь SPM (фиг. 4B), а гомозиготное ухо инициировало шесть SPM (фиг. . 4С). Позже, когда нормальные SPM инициируют два SM (рис. 4D), некоторые из SPM у мутантов Sos1 / + инициировали два SM (рис. 4E), в то время как очень немногие из SPM у мутантов Sos1 / Sos1 инициировали два SM. (Рисунок.4F). Вместо этого SPMs конвертируются непосредственно в SM, что подтверждается образованием чешуек (Fig. 4F).
Производство меньшего количества SPM могло произойти из-за первичного дефекта в размере IM. Таким образом, высота и ширина IM были измерены в Sos1 и нормальных ушах с помощью анализа SEM (рис. 3, D – F). Это показало, что Sos1 / Sos1 IM были на 80% шире нормальных соцветий, хотя разница была на грани статистической значимости (рис. 3B; P = 0,059). Кроме того, ИМ Sos1 / Sos1 составляли в среднем 71% роста нормальных ИМ, и это различие было статистически значимым (рис.3С; P = 0,014). Sos1 / + IM были промежуточными по размеру между нормальными и мутантами Sos1 / Sos1 . Следовательно, мутанты Sos1 продуцируют меньше SPM, предположительно из-за первичного дефекта в размере IM.
Таким образом, гистологический анализ и анализ SEM показывают, что мутанты Sos1 и продуцируют меньше колосков по двум причинам: (1) количество рядов SPM меньше; и (2) SPM непосредственно преобразуются в SM, не инициируя второй SM, что приводит к образованию одиночных, а не парных колосков.
Sos1 Мутация — антиморф
Анализ фенотипа Sos1 показал, что растения, гомозиготные по мутации Sos1 , имели более тяжелый фенотип, чем растения, гетерозиготные по Sos1 , и, следовательно, мутация является полудоминантной. Были определены четыре типа доминантных мутаций (Muller, 1932). Двумя типами мутаций с усилением функции являются мутации неоморфа, которые придают новую функцию, и мутации гиперморфа, которые вызывают повышенную экспрессию гена.Два типа мутаций с потерей функции — это гипоморф и антиморф. Мутации гипоморфа также называют гапло-недостаточными или чувствительными к дозе мутациями, потому что одной копии гена дикого типа недостаточно для функционирования. У антиморфов, которые также называют доминантно-негативными мутациями, мутантная копия гена нарушает функцию гена дикого типа (Veitia, 2007).
Чтобы различать эти типы мутаций, используется анализ доз, чтобы варьировать дозу копии гена дикого типа в мутантном фоне (Greene and Hake, 1994).У неоморфа не было бы эффекта варьирования дозы (Freeling and Hake, 1985; Poethig, 1988), в то время как у гиперморфа увеличение количества копий дикого типа привело бы к более серьезному фенотипу (Kessler et al. , 2002). С другой стороны, у гипо- и антиморфов добавление копий дикого типа будет вызывать ослабление фенотипа (Poethig, 1988; Nelson et al., 2002). Гипоморфа можно отличить от антиморфа, глядя на эффект изменения числа копий гена дикого типа на фоне дикого типа.Удаление одной копии гена дикого типа вызовет видимый фенотип, если ген был гапло-недостаточным и мутация была гипоморфной, но не если мутация была антиморфной.
Для определения фенотипического эффекта варьирования дозы sos1 дикого типа пыльцу гиперплоидов линии транслокации B-A, TB-4Sa, скрещивали с нормальными растениями или растениями, гетерозиготными по Sos1 . Скрещивание F 1 анализировали на уровень плоидности (путем оценки прерывания пыльцы; см. «Материалы и методы») и на серьезность фенотипа (путем подсчета процента одиночных колосков в кисточке и количества ядер в ухе. ).Эти результаты показали, что Sos1 растений с дополнительной копией короткого плеча хромосомы 4 ( Sos1 / + / + , гиперплоид) имели более слабый фенотип, чем растения, у которых отсутствовала нормальная копия короткого плеча хромосомы 4 ( Sos1 / — , гипоплоидный), поскольку у них был более высокий процент парных колосков (Рис. 5A
Рис. 5.
Анализ дозировки. A, Процент парных (серая полоса) по сравнению с одиночными (белыми) колосками в Sos1 растений, содержащих две копии хромосомы 4S дикого типа (гиперплоид) или без короткого плеча хромосомы 4 (гипоплоид).B. Процент парных (серых) по сравнению с одиночными (белыми) колосками в нормальных растениях, содержащих три копии хромосомы 4S дикого типа (гиперплоид) или одну копию хромосомы 4S дикого типа (гипоплоид).
Рисунок 5.
Анализ дозировки. A: процент парных (серая полоса) по сравнению с одиночными (белыми) колосками у растений Sos1 , содержащих две копии хромосомы 4S дикого типа (гиперплоид) или отсутствующих короткое плечо хромосомы 4 (гипоплоид). B. Процент парных (серых) по сравнению с одиночными (белыми) колосками в нормальных растениях, содержащих три копии хромосомы 4S дикого типа (гиперплоид) или одну копию хромосомы 4S дикого типа (гипоплоид).
). Аналогичные результаты наблюдались и в ухе (данные не показаны). Эти результаты показывают, что Sos1 не является неоморфом, так как имел место эффект варьирования дозировки гена, а не гиперморф, поскольку гиперплоид не был более серьезным, чем гипоплоид.
Чтобы определить, был ли Sos1 гипоморфной или антиморфной мутацией, мы создали серию доз для хромосомы 4S на генетическом фоне дикого типа. Нормальные растения, у которых отсутствует одна копия короткого плеча хромосомы 4, не имеют фенотипа Sos1 .Как в гипоплоиде (+/-), так и в гиперплоиде (+ / + / +) были в основном парные колоски (рис. 5B). Этот результат указывает на то, что ген sos1 дикого типа не является гапло-недостаточным и, следовательно, мутация Sos1 не является гипоморфом. Таким образом, мутация Sos1 , вероятно, является антиморфной или доминантно-отрицательной мутацией.
Sos1 Подавляет ra1 Мутантный фенотип
Sos1 мутанты имеют одиночные колоски вместо парных, потому что SPM продуцируют один SM вместо двух.Следовательно, SPM более детерминирован, чем обычно, у мутантов Sos1 . Следовательно, ген sos1 можно рассматривать как негативный регулятор детерминированности SPM. У мутантов ra1 SPM являются неопределенными, что приводит к образованию сильно разветвленных кисточек и ушей (Рис. 6, A и C
Рис. 6.
Генетическое взаимодействие между Sos1 и ra1 . A, Фотография зрелая кисточка всех генетических классов семейства F 2 , разделяющая на Sos1 и ra1 .B. Классификация количества различных типов подмышечных структур, создаваемых кисточкой. C. Фотография зрелых ушей всех генетических классов семейства F 2 , сегрегированных на Sos1 и ra1 . D к G, СЭМ соцветия початка. D, нормальное ухо, показывающее ряды парных SM. E, ra1 ухо, которое сильно разветвлено, так как все SPM преобразованы в ответвления. Ф, Сос1 / +; ra1 / ra1 ухо, в котором SPM производят дополнительные SM, но не так много, как ra1 .Г, Сос1 / Сос1; ra1 / ra1 колос, в котором SPM в основном дают одиночные колоски. На одной стороне наконечника нет SPM (стрелка). Масштабная линейка = 100 мкм м.
Рисунок 6.
Генетическое взаимодействие между Sos1 и ra1 . A, фотография зрелой кисточки всех генетических классов семейства F 2 , разделяющих на Sos1 и ra1 . B. Классификация количества различных типов подмышечных структур, создаваемых кисточкой.C. Фотография зрелых ушей всех генетических классов семейства F 2 , сегрегированных на Sos1 и ra1 . D к G, СЭМ соцветия початка. D, нормальное ухо, показывающее ряды парных SM. E, ra1 ухо, которое сильно разветвлено, так как все SPM преобразованы в ответвления. Ф, Сос1 / +; ra1 / ra1 ухо, в котором SPM производят дополнительные SM, но не так много, как ra1 . Г, Сос1 / Сос1; ra1 / ra1 колос, в котором SPM в основном дают одиночные колоски. На одной стороне наконечника нет SPM (стрелка).Масштабная линейка = 100 мкм м.
; Гернарт, 1912; Vollbrecht et al., 2005). Ген ra1 интерпретируется как положительный регулятор детерминированности SPM (Vollbrecht et al., 2005). Поскольку мутанты Sos1 и ra1 имеют противоположные эффекты на детерминированность SPM, мы проверили, действуют ли соответствующие гены по одному и тому же пути, создав двойные мутанты.
В кисточке, Сос1 / Сос1; Двойные мутанты ra1 / ra1 были менее разветвленными, чем одиночные мутанты ra1 (рис.6А). Чтобы количественно оценить эффекты обеих мутаций, ветви на кистях были удалены, классифицированы и подсчитаны с использованием схемы, аналогичной той, которая использовалась для анализа ra1 (рис. 6B; Vollbrecht et al., 2005). Нормальные растения производят ветви, прежде чем резко переключиться на производство пар колосков. ra1 Мутанты продуцируют несколько промежуточных продуктов между ветвями и парами колосков (Vollbrecht et al., 2005). Эти промежуточные продукты включают «смешанные ветви», которые представляют собой ветви со смесью одиночных и парных колосков; «Колосковые мультимеры» — ветви с одиночными колосками; и «тройные колоски».Наш анализ подтвердил, что, как и в предыдущем отчете (Vollbrecht et al., 2005), мутанты ra1 имели градацию фенотипа от ветвей до пар колосков с отсроченным переключением на идентичность SPM по сравнению с нормальным (рис. 6B). . Было обнаружено, что ветви у мутантов Sos1 представляют собой смешанные ветви, а не истинные ветви (Рис. 6B). Sos1 мутанты продуцировали меньше типов ветвей в целом и, следовательно, имели более ранний переход к идентичности SPM, чем обычно. В Сос1 / Сос1; ra1 / ra1 двойных мутантов, наблюдалось подавление фенотипа ra1 .Ветви были заменены смешанными ветвями, и было меньше всех типов ветвей по сравнению с одиночными мутантами ra1 и (рис. 6В).
Уменьшение количества типов филиалов в Сос1 / Сос1; Двойные мутанты ra1 / ra1 потенциально могут быть объяснены тем фактом, что мутанты Sos1 продуцируют меньше SPM (Fig. 3A). Итак, мы также оценили процент типов ветвей по сравнению с общим количеством подмышечных структур, создаваемых кисточкой. В этом случае мутация Sos1 все еще оказывала подавляющее действие на фенотип ra1 , поскольку процент всех типов ветвей уменьшился с 41.8% у одиночных мутантов ra1 до 18,7% у Sos1 / Sos1; ra1 / ra1 двойных мутантов. Следовательно, даже с учетом продукции меньшего количества SPM мутантами Sos1 , ветвление в двойной мутантной метелке подавлялось.
Подавление фенотипа ra1 Sos1 было даже более очевидным в ухе, чем в кисточке. Уши растений, которые были Sos1 / +; ra1 / ra1 были менее разветвленными, чем ra1 , и даже инициировали некоторые жизнеспособные ядра, что случается очень редко у одиночных мутантов ra1 .Кроме того, в Сос1 / Сос1; У двойного мутанта ra1 / ra1 ветвление было почти полностью подавлено. В самых крайних примерах уши выглядели как Sos1 , за исключением того, что они были меньше и более бесплодны (рис. 6C).
Чтобы исследовать онтогенетическую основу супрессии ra1 Sos1 , SEM-анализ был выполнен на ушах семей, сегрегированных по обеим мутациям. В нормальных ушах SPM производит два SM (рис. 6D), в то время как в ra1 ушах каждый SPM становится неопределенным и разветвляется, чтобы непрерывно производить SM повторяющимся образом (рис.6E). Сос1 / +; ra1 / ra1 уши были подавлены по сравнению с ra1 (рис. 6, E и F). Кончик напоминал Sos1 , но у основания уха SPM разветвлялся, чтобы продуцировать множественные SM, хотя и не так много, как у мутантов ra1 (Fig. 6F). В тяжелых случаях Сос1 / Сос1; ra1 / ra1 произвел уши, которые напоминали Sos1 (рис. 6G). Более того, у двойных мутантов иногда наблюдалось бесплодное пятно вдоль стороны уха (стрелка, фиг. 6G), фенотип, который не наблюдался ни у одного из одиночных мутантов.Этот бесплодный фенотип также был виден в зрелом ухе (рис. 6С). Следовательно, мутация Sos1 подавляла фенотип мутации ra1 как в кисточке, так и в ухе.
сос1; ra2 Двойные мутанты обладают синергетическим фенотипом
ra2 также работает в детерминировании SPM. У мутантов ra2 SPM становится неопределенным и дает дополнительные ветви и колоски в кисточке (Рис. 7A
Рис. 7.
Генетическое взаимодействие между Sos1 и ra2 . A, фотографии зрелой кисточки всех генетических классов семейства F 2 , разделяющего на Sos1 и ra2 . B. Классификация количества различных типов подмышечных структур, создаваемых кисточкой. C, вид крупным планом отдельных ветвей, произведенных в Sos1 / +; ra2 / ra2 кисточки двойных мутантов. D, вид крупным планом абортированных колосков, произведенных в Sos1 / Sos1; ra2 / ra2 кисточки двойных мутантов.E, фотография зрелого уха всех генетических классов семейства F 2 , разделяющего на Sos1 и ra2 . F, вид крупным планом подмышечных структур, образовавшихся в ушах семейства двойных мутантов. Нормальный и ra2 дают два ядра в ряду, мутанты Sos1 дают одно ядро в ряду, Sos / +; ra2 / ra2 производят длинные ответвления с несколькими ядрами, Sos1 / Sos1; ra2 / ra2 дает длинные стерильные ветки без ядер. G к J, СЭМ соцветия колоса.G — Нормальное ухо, показывающее ряды парных SM. H, ra2 ухо, в котором SPM образуют три SM или случайные ответвления. И, Сос1 / +; ra2 / ra2 колос, в котором СПМ образуют удлиненные ответвления. J, Sos1 / Sos1; ra2 / ra2 колос, в котором SPM образуют бесплодные удлиненные ветви. Масштабная линейка = 100 мкм м.
Рисунок 7.
Генетическое взаимодействие между Sos1 и ra2 . A, фотографии зрелой кисточки всех генетических классов семейства F 2 , разделяющего на Sos1 и ra2 .B. Классификация количества различных типов подмышечных структур, создаваемых кисточкой. C, вид крупным планом отдельных ветвей, произведенных в Sos1 / +; ra2 / ra2 кисточки двойных мутантов. D, вид крупным планом абортированных колосков, произведенных в Sos1 / Sos1; ra2 / ra2 кисточки двойных мутантов. E, фотография зрелого уха всех генетических классов семейства F 2 , разделяющего на Sos1 и ra2 . F, вид крупным планом подмышечных структур, образовавшихся в ушах семейства двойных мутантов.Нормальный и ra2 дают два ядра в ряду, мутанты Sos1 дают одно ядро в ряду, Sos / +; ra2 / ra2 производят длинные ответвления с несколькими ядрами, Sos1 / Sos1; ra2 / ra2 дает длинные стерильные ветки без ядер. G к J, СЭМ соцветия колоса. G — Нормальное ухо, показывающее ряды парных SM. H, ra2 ухо, в котором SPM образуют три SM или случайные ответвления. И, Сос1 / +; ra2 / ra2 колос, в котором СПМ образуют удлиненные ответвления. J, Sos1 / Sos1; ra2 / ra2 колос, в котором SPM образуют бесплодные удлиненные ветви.Масштабная линейка = 100 мкм м.
; Кемптон, 1923 г .; Никерсон и Дейл, 1955; Bortiri et al., 2006a). Однако ухо поражено лишь незначительно из-за нерегулярной гребли и случайных ветвей (рис. 7E). ra2 также играет уникальную роль в развитии соцветий, так как, в отличие от ra1 , ветви кисточки вертикальные, а ножки колосков удлиненные у мутантов ra2 (Bortiri et al., 2006a). Предполагается, что ra2 действует перед ra1 (Vollbrecht et al., 2005; Bortiri et al., 2006a).
Чтобы проверить, действует ли sos1 по тому же пути, что и ra2 , были сконструированы двойные мутанты. Удивительно, но Sos1 имел синергетическое взаимодействие с ra2 как в кисточке, так и в ухе (фиг. 7). Чтобы определить влияние мутации Sos1 на фенотип ra2 в кисточке, ветви были удалены, классифицированы и количественно определены с использованием той же системы классификации, которая использовалась ранее для ra1 и ra2 (Vollbrecht et al., 2005; Bortiri et al., 2006a). Мутанты ra2 , такие как ra1 , продуцировали промежуточные звенья между ветвями и колосками вдоль основного колоса, за исключением того, что фенотип был слабее, чем у ra1 (рис. 7B; Vollbrecht et al., 2005; Bortiri et al., 2006a). Оба одиночных мутанта Sos1 и ra2 имели небольшой эффект на общее количество подмышечных структур, продуцируемых кисточкой (Рис. 7B; Bortiri et al., 2006a). Поразительно, но было резкое уменьшение (более чем в 3 раза) общего количества подмышечных структур, производимых Sos1 / Sos1; ra2 / ra2 двойных мутантов по сравнению с любым одиночным мутантом (рис.7Б). Одиночные мутанты Sos1 и ra2 дают небольшое количество рудиментарных колосков с 1-2 чешуей без цветков (так называемые прерванные колоски). Подмышечные структуры, произведенные в Сос1 / Сос1; Двойные мутанты ra2 / ra2 почти полностью состояли из абортированных колосков (рис. 7, B и D). Мы интерпретируем эти фенотипы как синергетическое усиление дефектов ra2 .
В Сос1 ; ra2 с двойным мутантным ухом, усиление фенотипа ra2 также было очевидным.Уши мутантов ra2 лишь изредка дают ответвления (Vollbrecht et al., 2005; Bortiri et al., 2006a). Однако Sos1 / +; ra2 / ra2 и Sos1 / Sos1; Уши ra2 / ra2 были гораздо более разветвленными, чем уши ra2 (рис. 7E). Сос1 / +; ra2 / ra2 произвел ответвления с несколькими ядрами на них, а Sos1 / Sos1; Ветви ra2 / ra2 были длинными и бесплодными (рис. 7F).
Чтобы понять основу развития повышенного ветвления и бесплодия Sos1; ra2 двойные мутантные уши, SEM-анализ проводили на развивающихся ушах.В нормальных ушах SPMs продуцировали два SM (рис. 7G), тогда как у мутанта ra2 SPM продуцировал более двух SM и имелись случайные ответвления (рис. 7H). Модель Sos1 / +; Мутанты ra2 / ra2 были более разветвленными, чем ra2 , причем каждый SPM ветвился несколько раз (фиг. 7I). Ветви были более вытянутыми, чем ra2 , и давали мало SM (рис. 7, H и I). В Сос1 / Сос1; ra2 / ra2 , фенотип был дополнительно усилен удлиненными ветвями вместо SPM и даже меньшим количеством SM (рис.7J). Следовательно, Sos1 усиливал фенотип мутанта ra2 как в кисточке, так и в ухе.
Сос1; ra3 Двойные мутанты имеют аддитивный фенотип
ra3 также действует в детерминированности меристемы, хотя фенотип слабее, чем ra1 и ra2 (Satoh-Nagasawa et al., 2006). В кисточке мутанты ra3 дают несколько очень длинных ветвей, но не имеют дефектов детерминированности, наблюдаемых у мутантов ra1 и ra2 (данные не показаны).В ухе мутанты ra3 имеют неправильные ряды и ветви у основания уха (Рис. 8B
Рис. 8.
Генетическое взаимодействие между Sos1 и ra3 . A – D, Фотографии зрелых ушей от a Sos1; ra3 сегрегирующее семейство. От E до H, SEM-анализ ушей сегрегационного семейства Sos1; ra3 . Масштабная шкала = 100 μ м.
Рисунок 8.
Генетическое взаимодействие между Sos1 и ra3 .От A до D, фотографии зрелых ушей из Sos1; ra3 сегрегационная семья. От E до H, SEM-анализ ушей от Sos1; ra3 сегрегационная семья. Масштабная линейка = 100 мкм м.
; Сато-Нагасава и др., 2006). Эти дефекты вызваны превращением SPM в ветви и SM, производящие дополнительные структуры, включая дополнительные FM (Fig. 8F; Satoh-Nagasawa et al., 2006). В Сос1 / +; ra3 / ra3 двойные мутантные уши, ветви образовывались у основания уха, а одиночные колоски образовывались на кончике уха (рис.8C). Кроме того, аналогично ra3 одиночным мутантам, Sos1 / +; Колоски ra3 / ra3 давали дополнительные FM (рис. 8G). Сос1 / Сос1; уши двойного мутанта ra3 / ra3 имели фенотип, сходный с фенотипом Sos1 / +; ra3 / ra3 , за исключением того, что фенотип Sos1 был очевиден на большей протяженности уха (рис. 8D). Анализ SEM показал, что неопределенный SPM был произведен в основании уха (рис. 8H). Таким образом, двойные мутанты имели в основном аддитивные дефекты с одиночными рядами колосков, вызванными мутацией Sos1 на кончике, и дополнительными структурами, вызванными мутацией ra3 у основания уха.
Экспрессия
ra1 , ra2 и ra3 в Sos1 Мутанты
Для дальнейшего тестирования роли sos1 в пути ramosa был протестирован относительный уровень экспрессии мРНК ra1 , ra2 и ra3 у мутантов Sos1 с использованием количественной обратной транскрипции в реальном времени (RT ) -ПЦР. Результаты показали, что уровни мРНК ra1 были снижены у мутантов Sos1 / Sos1 (рис.9A
Фиг. 9.
Экспрессия ra1 , ra2 и ra3 в мутантах Sos1 и модель роли sos1 в детерминированности SPM. От A до C, эксперименты с ОТ-ПЦР в реальном времени, показывающие относительный уровень экспрессии (A) ra1 , (B) ra2 и (C) ra3 в Sos / + и Sos / Sos1 мутанты. C. Модель взаимодействия между генами Sos1 и ramosa . ra1 , ra2 и ra3 необходимы для обеспечения детерминированности SPM. Мы предполагаем, что Sos1 является отрицательным регулятором детерминированности SPM. ra2 и ra3 действуют перед ra1 , а также имеют роли, независимые от ra1 . Ас Сос1; Двойные мутанты ra1 напоминают одинарные мутанты Sos1 , sos1 располагается после ra1 . Ас Сос1; двойные мутанты ra2 обладают улучшенным фенотипом, подобным ra1; Двойные мутанты ra2 , sos1 , как предполагается, положительно регулируют ra1 . Sos1; Двойные мутанты ra3 обладают аддитивным фенотипом, что указывает на то, что ra3 действует независимым путем. Модель подтверждается исследованиями экспрессии, показывающими, что ra1 снижено, но ra2 и ra3 не изменились в мутантах Sos1 .
Фигура 9.
Экспрессия ra1 , ra2 и ra3 в мутантах Sos1 и модель роли sos1 в детерминированности SPM.От A до C, эксперименты с ОТ-ПЦР в реальном времени, показывающие относительный уровень экспрессии (A) ra1 , (B) ra2 и (C) ra3 в Sos / + и Sos / Sos1 мутанты. C. Модель взаимодействия между генами Sos1 и ramosa . ra1 , ra2 и ra3 необходимы для обеспечения детерминированности SPM. Мы предполагаем, что Sos1 является отрицательным регулятором детерминированности SPM. ra2 и ra3 действуют перед ra1 , а также имеют роли, независимые от ra1 .Ас Сос1; Двойные мутанты ra1 напоминают одинарные мутанты Sos1 , sos1 располагается после ra1 . Ас Сос1; двойные мутанты ra2 обладают улучшенным фенотипом, подобным ra1; Двойные мутанты ra2 , sos1 , как предполагается, положительно регулируют ra1 . Sos1; Двойные мутанты ra3 обладают аддитивным фенотипом, что указывает на то, что ra3 действует независимым путем. Модель подтверждается исследованиями экспрессии, показывающими, что ra1 снижено, но ra2 и ra3 не изменились в мутантах Sos1 .
). Поскольку уровни мРНК ra1 регулируются ra2 и ra3 (Vollbrecht et al., 2005; Satoh-Nagasawa et al., 2006), мы проверили, было ли снижение экспрессии ra1 из-за дефекта в ra2 или ra3 выражение. Однако уровни мРНК ra2 и ra3 не были затронуты у мутантов Sos1 / Sos1 (фиг. 9, B и C). Следовательно, мутация Sos1 специфически влияла на уровни экспрессии ra1 .
ОБСУЖДЕНИЕ
Самый поразительный дефект у мутантов Sos1 состоит в том, что SPM инициирует один, а не два SM. Кроме того, мутанты Sos1 имеют дефекты в инициации SPM, что, как мы предполагаем, связано с общим уменьшением апикального размера IM. Мутанты Sos1 также дают меньше ответвлений. Однако получаемые ветви не являются нормальными, а представляют собой смешанные ветви, которые более детерминированы, чем нормальные. Как только SM продуцируются мутантами Sos1 , они обычно дают два цветочка.Однако небольшой процент прерванных колосков (колосков без цветков) проявляется позже в развитии. О рудиментарных колосках также сообщили Doebley et al. (1995). Следовательно, мутация Sos1 и влияет на детерминированность всех меристем, образующихся во время развития соцветий.
Роль
sos1 в ramosa Путь к определению меристем
ra1 и ra2 мутанты сильно разветвлены, поскольку SPM являются неопределенными.Следовательно, роль генов ra1 и ra2 заключается в том, чтобы налагать детерминированность на SPM (Vollbrecht et al., 2005; Bortiri et al., 2006a), представленные сплошной стрелкой в модели, показанной на рисунке 9D. ra2 предположительно действует выше ra1 , потому что уровни мРНК ra1 снижены у мутантов ra2 , а двойные мутанты между ra2 и слабым аллелем ra1 имеют усиленный фенотип (Vollbrecht et al. ., 2005; Bortiri et al., 2006a; Сато-Нагасава и др., 2006). Кроме того, предполагается, что ra2 выполняет дополнительные роли, независимые от ra1 (Bortiri et al., 2006a).
У мутантов Sos1 SPM более детерминированы, чем обычно, производя один SM вместо двух. Мутация Sos1 является антиморфом, который представляет собой тип доминантной мутации потери функции (точнее, антагонист функции дикого типа). Следовательно, у мутантов Sos1 отсутствие нормальной функции гена sos1 вызывает повышение детерминированности.Одна интерпретация функции дикого типа гена sos1 заключается в противопоставлении детерминированности SPM, представленной полосой на фиг. 9D. Другой способ описания этого состоит в том, что ген sos1 придает неопределенность SPM, но поскольку SPM обычно является детерминированным, мы предполагаем, что sos1 ингибирует детерминированность. Поскольку ген sos1 ингибирует детерминированность SPM, в то время как гены ramosa способствуют детерминированности SPM, были сконструированы двойные мутанты для проверки генетического взаимодействия между генами sos1 и ramosa .Удивительно, но мы обнаружили разницу во взаимодействии между sos1 и ra1 , ra2 и ra3 .
Мы предлагаем модель, показанную на рисунке 9D, для учета всех данных генетического взаимодействия и экспрессии. Sos1; Двойные мутанты ra1 напоминают одиночные мутанты Sos1 . Поскольку одиночные мутанты Sos1 и ra1 имеют противоположные фенотипы, мы интерпретируем результат двойного мутанта как означающий, что ген sos1 дикого типа функционирует ниже ra1 (рис.9D; Эйвери и Вассерман, 1992). Следовательно, ra1 может придавать детерминированность SPM путем отрицательной регуляции гена sos1 . Неожиданно, Сос1; Двойные мутанты ra2 имели улучшенный фенотип по сравнению с одиночными мутантами ra2 . Sos1; Двойной мутант ra2 поразительно похож на двойной мутант между ra2 и слабым аллелем ra1 (Vollbrecht et al., 2005). Одна из гипотез, объясняющих усиление фенотипа ra2 мутацией Sos1 , была бы, если бы ген sos1 положительно регулировал ген ra1 .В поддержку этой гипотезы, уровни мРНК ra1 снижены у мутантов Sos1 . Следовательно, мы предполагаем, что sos1 действует в ветви ra1 пути ramosa , обеспечивая петлю отрицательной обратной связи для управления детерминированностью SPM (рис. 9D).
ra3 также предположительно действует выше ra1 , хотя, поскольку ra3 кодирует трегалозо-6-фосфатфосфатазу, это взаимодействие не может быть прямым (Satoh-Nagasawa et al., 2006). ra3 мутанты имеют слабые дефекты в детерминированности SPM, указывая тем самым, что ra3 также играет роль в детерминированности SPM (Satoh-Nagasawa et al., 2006). Sos1; Двойные мутанты ra3 имеют в основном аддитивные дефекты, подразумевая, что sos1 и ra3 действуют независимыми путями.
Мутация Sos1 оказала более подавляющее действие на фенотип ra1 в ухе, чем кисточка. Альтернативная интерпретация фенотипа метелки Sos1; ra1 будет заключаться в том, что мутации Sos1 и ra1 имеют аддитивный эффект, что указывает на то, что sos1 действует независимо от пути ra1 .Однако снижение экспрессии ra1 в мутантах Sos1 не подтверждает независимого взаимодействия. Более того, поскольку иногда наблюдается полное подавление фенотипа ra1 в Sos1; ra1 уха, мы поддерживаем гипотезу о том, что разница между фенотипами двойных мутантов метелки и уха обусловлена различиями в модифицирующих факторах между кисточкой и ухом. Различия в выраженности фенотипов метелки и колоса обычны для мутантов по детерминантности соцветий кукурузы (Irish, 1997a, 1997b; Laudencia-Chingcuanco and Hake, 2002; Kaplinsky and Freeling, 2003; Bortiri et al., 2006а). Одно из объяснений состоит в том, что кисточка формируется в апикальном положении, а ухо — в подмышечном, поэтому их гормональная среда, вероятно, различается. Кроме того, была продемонстрирована субфункционализация повторяющихся генов между кисточкой и ухом (Mena et al., 1996). Кроме того, степень ветвления зависит от условий окружающей среды, что указывает на то, что дополнительные модифицирующие факторы могут влиять на ветвление.
Сос1; Двойной мутант ra2 оказывал синергетическое действие на ветвление в ухе.Альтернативная интерпретация заключается в том, что Sos1; Фенотип двойного мутанта ra2 можно считать аддитивным, если, например, мутация ra2 привела к тому, что SPM стал неопределенным, но SPM не смог инициировать сидячие SM из-за мутации Sos1 . Однако, как SPM в Sos1; Двойные мутанты ra2 более неопределенны, чем одиночные мутанты ra2 , мы пришли к выводу, что взаимодействие между Sos1 и ra2 не аддитивно в ухе.Кроме того, влияние мутации Sos1 на фенотип ra2 в метелке не является аддитивным.
Таким образом, хотя можно представить себе и другие интерпретации, мы отдаем предпочтение модели, представленной на рисунке 9D, потому что она объясняет все фенотипы одиночных и двойных мутантов, а также исследования экспрессии. Доминирующие отрицательные мутации могут быть вызваны мутациями факторов транскрипции, которые, например, могут димеризовать, но не связывать ДНК, или могут связывать ДНК, но не активировать транскрипцию (Veitia, 2007).Основываясь на фенотипе мутанта Sos1 , генетических взаимодействиях с мутантами ra и снижении экспрессии ra1 у мутантов Sos1 , мы предполагаем, что sos1 может кодировать фактор транскрипции, который взаимодействует с ra1 или ra2 .
Дополнительные роли
sos1 и ramosa генов в развитии соцветий
Помимо влияния на детерминированность SPM, Sos1; ra1 и Sos; Двойные мутанты ra2 имели дополнительные дефекты развития соцветий.Это означает, что соответствующие гены играют дополнительные роли в развитии, которые ранее не были обнаружены. Sos1; Двойные мутанты ra1 давали бесплодные пятна в ухе, которые не наблюдались ни у одного из одиночных мутантов. Это синергетическое взаимодействие можно объяснить функцией обоих генов в SPM. Поскольку ra1 и ra2 не выражаются в апикальном IM, мы предполагаем, что это связано с дефектом самого SPM. Мы пришли к выводу, что, поскольку гены выполняют противоположные функции в SPM, в их отсутствие SPM иногда не запускается.Этот эффект был также замечен в кисточке, поскольку было общее сокращение количества подмышечных структур, произведенных в Sos1; двойной мутант ra1 . В Сос1; ra2 , было еще более серьезное снижение количества подмышечных структур, образующихся в кисточке. Эти результаты показывают, что гены sos1 , ra1 и ra2 играют перекрывающиеся роли в продукции SPM как в кисточке, так и в ухе.
В единичных мутантах Sos1 , ra1 и ra2 небольшое количество абортированных колосков образовывалось в кисточке, причем мутанты ra2 имели наиболее сильный эффект.В Сос1; ra1 , было несколько аддитивное увеличение количества абортированных колосков в кисточке по сравнению с любым одним мутантом. Однако в Сос1; ra2 , было синергетическое увеличение количества абортированных колосков. Фактически, почти все колоски, образовавшиеся в кисточке с двойным мутантом, были абортированы. Поэтому мы предлагаем, чтобы sos1 , ra1 и ra2 также работали в SM. Однако SMs все еще не определены в двойных мутантах Sos1; ra3 , сходных с одиночными мутантами ra3 , что указывает на то, что роль sos1 не зависит от ra3 в SM.
Роль
sos1 в регулировании размера IM
Мутанты Sos1 имеют меньший апикальный IM и меньшее количество SPM. Мы утверждаем, что дефекты в инициировании SPM происходят из-за дефектов в размере IM, поскольку есть другие примеры, когда увеличение или уменьшение размера меристемы влияло на инициирование SPM. Например, узловатых1 ( kn1 ) мутантов с потерей функции, которые также производят меньше ветвей и колосков в соцветии, уменьшили размер SAM в некоторых генетических фонах, хотя разницы в размере IM не было. продемонстрировано (Kerstetter et al., 1997; Vollbrecht et al., 2000). Противоположный фенотип наблюдается у мутантов с толстой кисточкой 1 ( td1 ) и фасциированных ушей2 ( fea2 ) мутантов, которые имеют более крупный IM и продуцируют больше SPM (Taguchi-Shiobara et al., 2001; Bommert et al. ., 2005). Интересно, что один из аллелей ra3 , ra3-fae1 , был первоначально идентифицирован как имеющий фасцитированный фенотип уха (Satoh-Nagasawa et al., 2006), что указывает на то, что путь ramosa может иметь косвенное влияние на IM . abphyl1 ( abph2 ) мутанты, которые влияют на филлотаксию листа, также имеют более крупную SAM (Jackson and Hake, 1999). abph2 кодирует регулятор цитокининового ответа, который негативно регулирует передачу сигналов цитокинина (Giulini et al., 2004). Кроме того, мутации, которые влияют на уровни цитокининов в рисе, также влияют на количество колосков (Ashikari et al., 2005; Kurakawa et al., 2007). Предполагается, что дефекты в мутантах kn1 , td1 и fea2 связаны с первичным дефектом уровней цитокининов (Barazesh and McSteen, 2008b), поскольку уровни цитокинина коррелируют с размером меристемы у eudicots (Sakakibara , 2006; Kyozuka, 2007).Имеют ли мутанты Sos1 дефекты в уровнях цитокининов или передаче сигналов, еще предстоит определить. Поскольку мутанты Sos1 и влияют на размер IM и детерминированность BM и SPM, мы предполагаем, что существует сходный основной дефект в поддержании или детерминированности как апикальных, так и подмышечных меристем.
Играет ли ауксин роль в функции
sos1 ?
Одиночные колоски на ножке также характерны для мутаций, влияющих на транспорт ауксина.Например, Бесплодное соцветие1 ( Bif1 ) и мутанты bif2 , которые дефектны в транспорте ауксина, продуцируют одиночные колоски (McSteen and Hake, 2001; McSteen et al., 2007; Barazesh and McSteen, 2008a). бесплодный стебель1 ( ba1 ), который, как предполагается, действует как перед транспортом ауксина, так и после него, также при мутации дает единичные колоски (Ritter et al., 2002; Gallavotti et al., 2004, 2008; Wu and McSteen, 2007; Скирпан и др., 2008). Более того, обработка нормальных соцветий кукурузы ингибиторами транспорта ауксина на более поздних этапах развития приводит к образованию единичных колосков (Wu and McSteen, 2007). Следовательно, транспорт ауксина необходим для инициирования сидячих колосков. Генетический анализ показывает Sos1; Двойные мутанты bif2 похожи на bif2 (дополнительный рисунок S1). Аналогично, ba1 является эпистатическим для Sos1 (дополнительный рисунок S1). Это можно интерпретировать как означающее, что sos1 действует по тому же пути, что и bif2 и ba1 , поскольку Sos1 , по-видимому, не усиливает фенотип bif2 или ba1 .Однако фенотипы bif2 и ba1 настолько серьезны, что было бы трудно увидеть улучшение. Таким образом, неясно, играет ли sos1 роль в обеспечении возможности ауксина, прямо или косвенно, инициировать сидячую SM.
В заключение, мы в настоящее время занимаемся точным картированием Sos1 для идентификации гена с помощью позиционного клонирования (Bortiri et al., 2006b). Клонирование гена sos1 позволит выяснить молекулярную основу его взаимодействия с путем ramosa .Более того, клонирование sos1 даст представление о его предполагаемой роли в эволюции парных колосков. Поскольку ra1 был идентифицирован только у Andropogoneae, было бы интересно определить, кооптировался ли sos1 в тот же момент времени во время диверсификации трав.
МАТЕРИАЛЫ И МЕТОДЫ
Рост растений и характеристика зрелого фенотипа
Аллель Sos1-Reference был получен из Центра сотрудничества генетики кукурузы (запас 427I) и шесть раз скрещен с генетическим фоном B73 кукурузы ( Zea mays ).Анализ зрелого фенотипа проводили на растениях, выращиваемых в Рок-Спрингс, штат Пенсильвания, летом. Растения были генотипированы как гетерозиготы или гомозиготы Sos1, с использованием SSR-маркера umc1294 , который является ближайшим генетическим маркером, идентифицированным на данный момент (umc1294F, 5′-GCC GTC AAC GGG CTT AAA CT-3 ‘и umc1294R, 5’-GCC TCC ACG TCT CTC GTC TCT T-3 ′). Для характеристики фенотипа кисточки и колосья собирали по достижении зрелости у нормальных растений Sos1 / + и Sos1 / Sos1 из сегрегированных семейств.Количество ветвей и количество парных колосков по сравнению с одиночными на главном колосе подсчитывали на кистях после цветения. Размер выборки составлял 28 нормальных, 35 Sos1 / + и 33 Sos1 / Sos1 в данных, представленных на рис. 1, B и C. Число ядер подсчитывали на открытых опыленных початках. Размер выборки был 16 нормальных, 13 Sos1 / + и 10 Sos1 / Sos1 в данных, представленных на рис. 1, E и F.
SEM и гистология
Незрелые колосья (5–20 мм) собирали с растений, выращиваемых в течение 8 недель летом в Рок-Спрингс, штат Пенсильвания.Незрелые кисточки (3–6 мм) получали от 5-недельных растений, выращенных весной в теплице с дополнительным освещением. И кисточки, и уши были закреплены и подготовлены для SEM, как описано (Wu and McSteen, 2007). Количество SPM по окружности апикального IM определялось путем установки верхней части уха на заглушке SEM и просмотра образцов сверху ( n = 11 нормальных, 8 Sos1 / + и 7 Sos1 / Сос1 ). Для измерения размера IM образцы колосов просматривали сбоку, и ширину IM измеряли поперек меристемы непосредственно над точкой, где были видны примордии прицветника, а высоту измеряли перпендикулярно от этой линии к верхушке соцветия ( n = 7 нормальных, 7 Sos1 / + и 6 Sos1 / Sos1 ).Кроме того, кисточки и ушки заливали воском и делали срезы для окрашивания ТВО, как описано (Wu and McSteen, 2007).
Анализ дозировки
Анализ дозировки проводили путем скрещивания известных гиперплоидов линии транслокации B-A, TB-4Sa (помеченных как сахаристый1 , Coop Stock 421D) как самцов с нормальными растениями или растениями, гетерозиготными по Sos1 . TB-4Sa — это реципрокная транслокация короткого плеча хромосомы 4 с сверхнормальной B-хромосомой, которая из-за очень высокой скорости нерасхождения во время второго митотического деления развития пыльцы дает пыльцу с двумя или нулевыми копиями хромосома 4S (Роман, 1947; Беккет, 1978, 1994).Плоидность потомства F 1 может быть определена по скорости аборта пыльцы (Роман, 1947; Карлсон, 1994). Аборт пыльцы оценивали с помощью карманного микроскопа по утрам подряд, когда пыльники начинали отделять свежую пыльцу (Phillips, 1994). Как наблюдалось ранее для TB4Sa, выкидыш пыльцы у гипоплоидных растений составлял 50%, а у гиперплоидных растений — от 2% до 10% (Carlson, 1994; D. Auger, Университет штата Южная Дакота, личное сообщение). Кисточки и уши гипоплоидов и гиперплоидов собирали по достижении зрелости для фенотипического анализа.Подсчитывали процент парных и одиночных колосков на 15-сантиметровом участке главного колоса кисточки после цветения. Для анализа, показанного на рисунке 4, размеры образцов составляли 13 Sos1 / + / + , 6 Sos1 / — , 8 + / + / + и 4 +/- кисточек.
Анализ двойных мутантов
Все семьи, разделяющие двойные мутанты, были созданы на генетическом фоне B73 и были высажены летом в Рок-Спрингс, штат Пенсильвания, в течение двух полевых сезонов. На каждую комбинацию двойных мутантов ежегодно высаживали не менее 360 растений.Все растения были генотипированы для Sos1 с маркером SSR umc1294 , как описано выше. Каждое семейство двойных мутантов оценивали, и анализ хи-квадрат не смог отклонить ожидаемый коэффициент сегрегации (данные не показаны).
Семьи сегрегационные по Сос1; bif2 и Sos1; ba1 были генотипированы по мутациям bif2 и ba1 , как описано (Skirpan et al., 2008). По мере созревания подсчитывали количество веточек кисточки и колосков на главном колосе.Для анализа, показанного на дополнительном рисунке S1B, размер выборки был 2 нормальных, 4 Sos1 / + , 11 Sos1 / Sos1 , 17 bif2 / bif2 , 13 Sos1 / +; bif2 / bif2 и 13 Сос1 / Сос1; Биф2 / Биф2 . Для анализа, показанного на дополнительном рисунке S1D, размер выборки был 2 нормальных, 5 Sos1 / + , 7 Sos1 / Sos1 , 11 ba1 / ba1 , 11 Sos1 / +; ba1 / ba1 и 8 Сос1 / Сос1; Ба1 / Ба1 .
Раздельные семьи Sos1; ra1 и Sos1; ra2 оценивали, а фенотип кисточки и ушей анализировали в зрелом возрасте (9 недель).Ветви были удалены с кисточек и классифицированы согласно Vollbrecht et al. (2005) и Bortiri et al. (2006a). Для анализа, показанного на рисунке 6B, размер выборки был 4 нормальных, 4 Sos1 / + , 3 Sos1 / Sos1 , 9 ra1 / ra1 , 10 Sos1 / +; ra1 / ra1 и 10 Sos1 / Sos1; Ра1 / Ра1 . Для анализа, показанного на рисунке 7B, размер выборки был 4 нормальных, 4 Sos1 / + , 6 Sos1 / Sos1 , 6 ra2 / ra2 , 8 Sos1 / +; ra2 / ra2 и 9 Sos1 / Sos1; Ра2 / Ра2 .Семьи сегрегационные по Сос1; ra3 оценивали только по фенотипу ушей, поскольку фенотип метелки ra3 был очень слабым.
На всех графиках представлено среднее значение ± стандартное отклонение от среднего. Значения вероятности были определены на основе двусторонних тестов Стьюдента t , выполненных в Microsoft Excel 2003.
ОТ-ПЦР в реальном времени
Тотальную РНК экстрагировали из кисточек размером 4-5 мм из нормальных, Sos1 / + и Sos1 / Sos1 растений с помощью набора NucleoSpin RNA Plant (Macherey-Nagel GmbH & Co.). От четырех до шести образцов из каждого класса использовали в качестве биологических повторений, выполняли три технических повторения для каждого биологического повторения, и весь эксперимент повторяли три раза. Синтез кДНК, ОТ-ПЦР в реальном времени и анализ выполняли, как описано (Barazesh and McSteen, 2008a). Для обнаружения экспрессии ra1 зонд Такмана был (FAM-5′-ATC CAC AGG CTG GAC AGG GCC A-3′-BHQ), а праймеры RT-PCR были ra1F (5′-GCT GGG AGG CCA CAT GAA- 3 ‘) и ra1R (5′-GTG AAG TGT ACT GTT GGT GGA TCA G-3’).Для обнаружения экспрессии ra2 зонд Такмана был (FAM-5′-TCG TCA TTA GTA GCT CCC CAG GCG C-3′-BHQ), а праймеры RT-PCR были ra2F (5′-TGC TAC TTC ATG CGG AAC CA-3 ‘) и ra2R (5′-TTA GCC ACG GAA GCG TAA GG-3’). Для обнаружения экспрессии ra3 зонд Такмана был (FAM-5′-CCG TCC ACT TCC GCT GCG TCC-3′-BHQ), а праймеры RT-PCR были ra3F (5′-CCA GGG TGG AGC ACA ACA A- 3 ‘) и ra3R (5’-CTC GTT CAC GGC ATT CCA T). Внутренний контроль для амплификации — убиквитин ( убкв ).Для обнаружения экспрессии ubq зонд Такмана был (5′-FAM-AAA TCC ACC CGT CGG CAC CTC C-3 ‘BHQ), а праймеры RT-PCR были ubqF (5’-CTC TTT CCC CAA CCT CGT GTT- 3 ‘) и ubqR (5′-ACG AGC GGC GTA CCT TGA-3’).
Дополнительные данные
В онлайн-версии статьи доступны следующие материалы.
БЛАГОДАРНОСТИ
Мы благодарим Эрика Фоллбрехта за указание на сходство между Sos1; ra2 и ra1; двойные мутанты ra2 , Дэвид Барнс и Сара Хейк за первое открытие Sos1; ra2 и двух анонимных рецензентов за полезные предложения.Мы благодарим Деба Гроув из Центра нуклеиновых кислот Института Хака за проведение экспериментов ОТ-ПЦР в реальном времени, Мисси Хейзен из Центра электронной микроскопии Института Хака за обучение работе с SEM, а также Джеффри Бутербо, Джейсона Хоара, Криса Кука, Кима Филлипса, Мэтту Дэвису и Крису Хадсону за помощь в полевых условиях и за генотипирование Sos1 . Мы благодарим Maize Coop за семена Sos1 , Фрэнка Бейкера за семена гиперплоидов TB4Sa и Дона Оже за советы по анализу дозировки туберкулеза. Благодарим Тони Омейса и Скотта Харккома за уход за растениями.Мы благодарим сотрудников лабораторий Макстина и Брауна за обсуждение и комментарии к рукописи.
ЦИТИРОВАННАЯ ЛИТЕРАТУРА
Асикари М., Сакакибара Х., Лин С.И., Ямамото Т., Такаши Т., Нишимура А., Анхелес Э.Р., Циан К., Китано Х., Мацуока М. (
2005
) Цитокининоксидаза регулирует производство зерна риса.
Наука
309
:
741
—
745
Эйвери Л., Вассерман С. (
1992
) Упорядочивающая функция гена: интерпретация эпистаза в регуляторных иерархиях.
Тенденции Genet
8
:
312
—
316
Баразеш С., Макстин П. (
2008
a) бесплодное соцветие1 участвует в органогенезе во время вегетативного развития и развития соцветий кукурузы.
Генетика
179
:
389
—
401
Баразеш С., Макстин П. (
2008
b) Гормональный контроль развития соцветий травы.
Trends Plant Sci
13
:
656
—
662
Beckett JB (
1978
) Транслокации B-A у кукурузы.I. Использование для определения местоположения генов по хромосомным плечам.
J Hered
69
:
27
—
36
Beckett JB (
1994
) Определение местоположения рецессивных генов в плече хромосомы с транслокациями B-A. В М. Фрилинг, В. Уолбот, ред., Справочник по кукурузе. Springer-Verlag, New York, pp 315–327
Bommert P, Lunde C, Nardmann J, Vollbrecht E, Running M, Jackson D, Hake S, Werr W (
2005
) толстый карлик с кисточкой1 кодирует предполагаемый ортолог кукурузы из семейства рецепторов, богатых лейцином CLAVATA1 арабидопсиса. киназа.
Развитие
132
:
1235
—
1245
Bortiri E, Chuck G, Vollbrecht E, Rocheford T, Martienssen R, Hake S (
2006
a) ramosa2 кодирует белок LATERAL ORGAN BOUNDARY domain, который определяет судьбу стволовых клеток в меристемах ветвей кукурузы.
Растительная ячейка
18
:
574
—
585
Bortiri E, Hake S (
2007
) Цветение и определенность кукурузы.
J Exp Bot
58
:
909
—
916
Бортири Э., Джексон Д., Хейк С. (
2006
b) Достижения в геномике кукурузы: появление позиционного клонирования.
Curr Opin Plant Biol
9
:
164
—
171
Carlson WR (
1994
) Манипуляция транслокацией B-A. В М. Фрилинг, В. Уолбот, ред., Справочник по кукурузе. Springer-Verlag, New York, pp 308–314
Cheng PC, Greyson RI, Walden DB (
1983
) Инициирование органа и развитие однополых цветков в кисточке и ухе Zea mays .
Am J Bot
70
:
450
—
462
Chuck G, Meeley RB, Hake S (
1998
) Контроль судьбы меристемы колосков кукурузы с помощью APETALA2 -подобного гена неопределенного колоска1 .
Genes Dev
12
:
1145
—
1154
Clifford HT (
1987
) Морфология колосков и цветков. В Т. Р. Содерстрем, К. В. Хилу, К. С. Кэмпбелл, М. Е. Баркворт, ред., «Систематика и эволюция травы».Smithsonian Institution Press, Вашингтон, округ Колумбия, стр. 21–30
Doebley J, Stec A, Kent B (
1995
) Супрессор сидячих колосков1 ( Sos1 ): доминантный мутант, влияющий на развитие соцветий кукурузы.
Am J Bot
82
:
571
—
577
Доуст А.Н., Келлогг Е.А. (
2002
) Диверсификация соцветий в кладе паникоидной «щетинистой травы» (Paniceae, Poaceae): данные молекулярной филогении и морфологии развития.
Am J Bot
89
:
1203
—
1222
Freeling M, Hake S (
1985
) Генетика развития мутантов, которые определяют узловатые листья кукурузы.
Генетика
11
:
617
—
634
Gallavotti A, Yang Y, Schmidt RJ, Jackson D (
2008
) Взаимосвязь между транспортом ауксина и ветвлением кукурузы.
Plant Physiol
147
:
1913
—
1923
Галлавотти А., Чжао К., Киозука Дж., Мили Р. Б., Риттер М., Добли Дж. Ф., Пе М. Е., Шмидт Р. Дж. (
2004
) Роль бесплодного стебля 1 в архитектуре кукурузы.
Природа
432
:
630
—
635
Gernart W (
1912
) Новый подвид Zea mays L.
Am Nat
46
:
616
—
622
Джулини А., Ван Дж., Джексон Д. (
2004
) Контроль филлотаксии с помощью цитокинин-индуцируемого гомолога регулятора ответа ABPHYL1.
Природа
430
:
1031
—
1034
Рабочая группа по филогении трав (
2001
) Филогения и подсемейная классификация трав ( Poaceae ).
Энн Мисс Бот Гард
88
:
373
—
457
Greene B, Hake S (
1994
) Использование сегментарных анеуплоидов для анализа мутантов. В М. Фрилинг, В. Уолбот, ред., Справочник по кукурузе. Springer-Verlag, New York, pp 270–273
Irish EE (
1997
a) Класс II семян метелки. Мутации свидетельствуют о множестве типов меристем соцветий у кукурузы (Poaceae).
Am J Bot
84
:
1502
—
1515
Irish EE (
1997
b) Экспериментальный анализ развития метелки у мутанта кукурузы Tassel seed 6 .
Plant Physiol
114
:
817
—
825
Irish EE (
1998
) Колоски травы: острая проблема.
Биологические исследования
20
:
789
—
793
Джексон Д., Хейк С. (
1999
) Контроль филлотаксии кукурузы с помощью гена abphyl1 .
Развитие
126
:
315
—
323
Каплинский Н.Дж., Фрилинг М. (
2003
) Комбинаторный контроль идентичности меристемы в соцветиях кукурузы.
Развитие
130
:
1149
—
1158
Kellogg EA (
2000
) Молекулярная и морфологическая эволюция Andropogoneae. В SWL Jacobs, JE Everett, eds, Grasses: Systematics and Evolution. CSIRO, Коллингвуд, Австралия, стр. 149–158
Kellogg EA (
2001
) История эволюции трав.
Plant Physiol
125
:
1198
—
1205
Kellogg EA (
2007
) Цветочные проявления: генетический контроль соцветий травы.
Curr Opin Plant Biol
10
:
26
—
31
Kempton JH (
1923
) Наследственные признаки кукурузы XIV: разветвленные колосья.
J Hered
14
:
243
—
252
Керстеттер Р.А., Лауденсия-Чингкуанко Д., Смит Л.Г., Хейк С. (
1997
) Мутации с потерей функции в гене гомеобокса кукурузы, узловатый1 , нарушают поддержание меристемы побегов.
Развитие
124
:
3045
—
3054
Kessler S, Seiki S, Sinha N (
2002
) Xcl1 вызывает отложенные косые периклинальные деления клеток в развивающихся листьях кукурузы, что приводит к дифференцировке клеток по клонам, а не по положению.
Развитие
129
:
1859
—
1869
Киссельбах Т.А. (
1949
) Структура и воспроизводство кукурузы. Лабораторная пресса Колд-Спринг-Харбор, Колд-Спринг-Харбор, Нью-Йорк, стр. 37–63
Куракава Т., Уэда Н., Маэкава М., Кобаяши К., Кодзима М., Нагато Ю., Сакакибара Х., Киозука Дж. (
2007
) Прямой контроль активности меристемы побегов с помощью фермента, активирующего цитокинин.
Природа
445
:
652
—
655
Kyozuka J (
2007
) Контроль функции меристемы побегов и корней цитокинином.
Curr Opin Plant Biol
10
:
442
—
446
Laudencia-Chingcuanco D, Hake S (
2002
) Неопределенный ген вершины цветка регулирует детерминированность и идентичность меристемы в соцветии кукурузы.
Развитие
129
:
2629
—
2638
McSteen P (
2006
) Ветвление: путь ramosa и эволюция морфологии соцветий травы.
Растительная ячейка
18
:
518
—
522
McSteen P, Hake S (
2001
) бесплодное соцветие2 регулирует развитие пазушной меристемы в соцветии кукурузы.
Развитие
128
:
2881
—
2891
Макстин П., Лауденсия-Чингкуанко Д., Коласанти Дж. (
2000
) Цветочек под любым другим названием: контроль идентичности меристемы у кукурузы.
Trends Plant Sci
5
:
61
—
66
McSteen P, Malcomber S, Skirpan A, Lunde C, Wu X, Kellogg E, Hake S (
2007
) бесплодное соцветие2 кодирует ко-ортолог серин / треониновой киназы PINOID и необходим для органогенеза во время соцветие и вегетативное развитие кукурузы.
Plant Physiol
144
:
1000
—
1011
Мена М., Амброуз Б.А., Мили Р.Б., Бриггс С.П., Янофски М.Ф., Шмидт Р.Дж. (
1996
) Диверсификация активности С-функции в развитии цветков кукурузы.
Наука
274
:
1537
—
1540
Muller HJ (
1932
) Дальнейшие исследования природы и причин генных мутаций. В DF Jones, WI Menasha, eds, Proceedings of the Six International Congress of Genetics.Бруклинский ботанический сад, Бруклин, Нью-Йорк, стр. 213–255
Nelson JM, Lane B, Freeling M (
2002
) Экспрессии мутантного гена кукурузы в вентральном эпидермисе листа достаточно, чтобы сигнализировать о переключении дорсовентральной оси листа.
Развитие
129
:
4581
—
4589
Nickerson NH, Dale EE (
1955
) Модификации кисточки в Zea mays .
Энн Мисс Бот Гард
42
:
195
—
211
Филлипс Р.Л. (
1994
) Классификация абортов пыльцой в полевых условиях. В М. Фрилинг, В. Уолбот, ред., Справочник по кукурузе. Springer-Verlag, New York, pp 297–298
Poethig RS (
1988
) Гетерохронные мутации, влияющие на развитие побегов кукурузы.
Генетика
119
:
959
—
973
Ritter MK, Padilla CM, Schmidt RJ (
2002
) Мутант кукурузы бесплодный стебель1 дефектен в развитии подмышечной меристемы.
Am J Bot
89
:
203
—
210
Роман Х (
1947
) Митотическое нерасхождение в случае обменов с участием хромосомы В-типа кукурузы.
Генетика
32
:
391
—
409
Сабловски Р. (
2007
) Цветение и определенность у Arabidopsis.
J Exp Bot
58
:
899
—
907
Сакакибара Н (
2006
) Цитокинины: активность, биосинтез и транслокация.
Анну Рев Завод Биол
57
:
431
—
449
Сато-Нагасава Н., Нагасава Н., Малкомбер С., Сакаи Х., Джексон Д. (
2006
) Фермент метаболизма трегалозы контролирует структуру соцветий кукурузы.
Природа
441
:
227
—
230
Skirpan A, Wu X, McSteen P (
2008
) Генетическое и физическое взаимодействие предполагает, что бесплодных стеблей1 является целью бесплодных соцветий2 в развитии соцветий кукурузы.
Завод J
55
:
81
—
91
Стивс Т., Сассекс I (
1989
) Модели в развитии растений. Издательство Кембриджского университета, Кембридж, Великобритания
Taguchi-Shiobara F, Yuan Z, Hake S, Jackson D (
2001
) Ген фасциированного уха кодирует рецептор-подобный белок с высоким содержанием лейцина, который регулирует пролиферацию меристемы побегов кукурузы.
Genes Dev
15
:
2755
—
2766
Veitia RA (
2007
) Изучение молекулярной этиологии доминантно-отрицательных мутаций.
Растительная ячейка
19
:
3843
—
3851
Vollbrecht E, Reiser L, Hake S (
2000
) Размер меристемы побега зависит от инбредного фона и присутствия гена гомеобокса кукурузы, с узлом1 .